Xantina oxidasa
La xantina oxidasa es una molibdoflavoenzima distribuida en diversas especies, desde las bacterias hasta los seres humanos y algunos tejidos en mamíferos. Su enfoque de estudio se debe a su afinidad por producir especies reactivas de oxígeno, fenómeno implicado en varios estados patológicos. Esta enzima se encuentra en dos formas, la Xantina Deshidrogenasa NAD+-dependiente, que produce NADH y urato, la cual puede ser transformado en Xantina Oxidasa que es dependiente del oxígeno, la cual origina el anión radical superóxido (O2) y peróxido de hidrógeno (H2O2) y urato.[2]
| Xantina oxidasa | ||||
|---|---|---|---|---|
 Estructura cristalográfica de la xantina oxidasa bovina.[1] | ||||
| Estructuras disponibles | ||||
| PDB |
Lista de códigos PDB 1FIQ
| |||
| Identificadores | ||||
| Símbolo | XDH (HGNC: 12805) | |||
| Identificadores externos |
Bases de datos de enzimas
| |||
| Número EC | 1.17.3.2 | |||
| Locus | Cr. 2 p23.1 | |||
| Ortólogos | ||||
| Especies |
| |||
| Entrez |
| |||
| UniProt |
| |||
| RefSeq (ARNm) |
| |||
| PubMed (Búsqueda) |
| |||
| PMC (Búsqueda) |
| |||
Reacción editar
Las siguientes reacciones químicas son catalizadas por la xantina oxidasa:
- hipoxantina + H2O + O2 xantina + H2O2
- xantina + H2O + O2 ácido úrico + H2O2

-
 Hipoxantina (un átomo de oxígeno)
Hipoxantina (un átomo de oxígeno) -
 Xantina (dos átomos de oxígeno)
Xantina (dos átomos de oxígeno) -
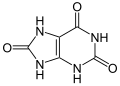 Ácido úrico (tres átomos de oxígeno)
Ácido úrico (tres átomos de oxígeno)



Estructura XOR editar
La estructura primaria de la XOR humana contiene una secuencia de 1335 residuos de aminoácidos y presenta 90 % de homología con otros mamíferos. Por otro lado, la estructura tridimensional se logró obtener a partir de los resultados del análisis de la estructura cristalina de otras enzimas, que al igual que la XOR presentan un cofactor molibdeno, tal es el caso del monóxido de carbono deshidrogenasa (CODH) presente en la bacteria aerobia Oligotropha carboxidovorans, la cual sirve como modelo para la XO, debido a su parecido en la presencia de: el cofactor molibdopterina (Mo-co), 2 centros (Fe2-S2) y un grupo FAD cómo centros redox. El dominio N-terminal de la XOR contiene 8 residuos de cisteína fuertemente conservados, los cuales sirven como ligandos para unir los centros (Fe2-S2), cada unión está conformada por 4 cisteínas, en donde la más lejana al grupo N-terminal presenta un plegamiento distinto, característica que diferencia esta enzima de las demás. Los centros Fe/S son llamados cómo Fe/S I y Fe/S II, con respecto a la distancia que los separa del grupo Mo-co. El centro Fe/S II se encuentra cerca de la superficie de la enzima, dicha distancia de la superficie le permite transferir electrones a su aceptor electrónico fisiológico, FAD, la distancia es de 8.7 A°. La estructura tridimensional de la XOR de leche bovina fue estudiada, encontrando que cada subunidad contiene 3 dominios y 2 segmentos peptídicos conectores, tales son:
- Dominio N-terminal que incluye los residuos de aminoácidos y contiene los sitios de unión para los centros (Fe2-S2). Este dominio se encuentra conformado desde el primer hasta el 165 aminoácido.
- Posteriormente se encuentra el segmento conector que incluye los residuos 166 al 225.
- Domino de unión al FAD con los residuos 226 al 531.
- Segmento conector, que incluye a los residuos 532 – 589.
- Dominio C-terminal para unir al Mo-co, incluye los residuos 590 en adelante.
Los potenciales REDOX de los diferentes centros garantizan que la transferencia electrónica sea termodinámicamente favorable, siendo el sentido de esta transferencia.[2] Mo-co Fe/S I Fe/S II FAD
Propiedades y características editar
Es una proteína compuesta por su porción apoproteica, que la conforman 2 cadenas polipeptídicas idénticas y 8 centros redox, es un homodímero de 300 Kd de peso molecular, cada subunidad tiene 4 centros redox, 1 molécula de molibdopterina tiene un átomo de molibdeno como cofactor, también tiene 2 centros ferro sulfurados (Fe2-S2) y contiene 1 molécula de dinucléotido de falvinadenina (FAD). Las dos formas de la enzima (XDH y XO) se pueden interconvertir con la excepción en las aves, ya que estás solo tienen la forma XDH.[2]
Genética editar
El gen de la XOR humana (hXOR) está en el brazo corto del cromosoma 2, 2p22, y su estructura ha sido dilucidada. Presenta 36 exones y su estructura exón-intrón está altamente conservada respecto a otras especies como el ratón y difiere significativamente de la estructura del gen en especies menos relacionadas como la Drosofila, cuya estructura es más compacta con solo 4-5 exones. La secuencia del cDNA humano ha sido reportada.[2]
Formas de la XOR editar
Previamente se han mencionado dos formas de la Xantina, tales son la Xantina Deshidrogenasa y la Xantina Oxidasa, el mecanismo para pasar de una a otra se explican a continuación:
Conversión reversible. Mediante este proceso la oxidación de grupos sulfhidrilos forman puentes disulfuros intrasubunitarios los cuales permiten la conversión de XDH a XO, mientras que la reducción de estos enlaces opera en sentido contrario. Esta conversión se comprobó con la cromatografía en gel, ya que la formación de puentes disulfuro reduce el volumen hidrodinámico de la enzima y con la electroforesis en gel bidimensional de la XO digerida con quimiotripsina se demostró la presencia de una heterogeneidad conformacional, medida por puentes disulfuros.
Conversión irreversible. Para esta conversión se lleva a cabo la proteólisis de la XOR, ambas formas de la XOR experimentan proteólisis limitada por tripsina y quimiotripsina, la primera separa ambas formas de la enzima en la lisina 184 mientras que la segunda divide la XDH en metionina 181, pero la XO la divide en metionina 181 y fenilalanina 560. La proteólisis quimiotríptica previene la conversión reductiva de XO en XDH. Además, los residuos de metionina 181 y lisina 184 están incluidos en un fragmento peptídico hidrofílico expuesto al solvente en ambas formas de la XOR. En contraste, la phe560 pertenece a una región hidrofóbica, por lo que ha sido hipotetizado que tenga poco acceso al solvente en condiciones fisiológicas, y por tanto, baja probabilidad de ser escindida por la quimiotripsina. La observación de que este sitio comienza a ser accesible a la quimiotripsina solamente luego de la conversión de XDH en XO, sugiere que la formación de puentes disulfuros produce cambios conformacionales dentro de la región que contiene a la phe560. Durante la conversión de XDH a XO, los principales cambios involucran al sitio de unión y el FAD, el cual tiene dos efectos importantes en cuanto al uso del oxígeno molecular cómo agente oxidante para la XO, los pasos se describen a continuación:
- El acceso al NAD+ a la superficie del anillo de isoaloxacina del FAD queda bloqueado.
- El entorno alrededor del FAD queda cargado positivamente, gracias al reemplazo del Asp429, desde su sitio de unión al anillo flavínico por la Arg426
A pesar de estos 2 efectos, el resto de la región del sitio de unión al FAD donde puede unirse el O2 es poco afectada por estos cambios.[2]
Funciones de la XO editar
La principal función de esta metaloenzima es hidroxilar una serie de sustratos, tales como hipoxantina, la cual pasa a xantina y esta a su vez en ácido úrico. Los demás productos serán dinucleótido de nicotinaminadenina reducido (NADH.H+) o (O2) y/o (H2O2) según haya actuado el NAD+ o el (O2) como agente reductor para la enzima respectivamente. Gracias a su capacidad generadora de especies reactiva se centra el interés en esta metaloenzima. También hidroxila otros sustratos como N-heterociclos y aldehídos. Ambas formas de la enzima pueden reducir el (O2) pero, la unión del NAD+ a la XDH inhibe la unión del (O2). Otra función de la xantina es que pueda fungir como una NADH oxidasa, la cual es capaz de donar sus electrones directamente al FAD. Se ha demostrado la capacidad de la enzima para catalizar la reducción de nitratos a nitritos, y de nitritos a óxido nítrico (NO). En estos casos, los nitratos son reducidos por el cofactor de molibdeno, mientras que nitritos son reducidos por el FAD. Liberación del hierro de la ferritina, que puede ocurrir por 2 mecanismos.[2]
- Mecanismo (O2)- dependiente, responsable del 80 % de esta separación.
- Mecanismo (O2)- independiente. Este efecto de la XO adquiere su importancia en el estudio de los tejidos dañados por la acción de las especies reactivas de oxígeno. Esto se debe a la toxicidad del radical superóxido en presencia de hierro libre, ya sea por la formación de radical hidroxilo, radiales ferrilos o radicales perferrilos.
Formas funcionales de la xantina oxidorreductasa editar
Forma activa XDH XO editar
Xantina Deshidrogenasa: representa 80 % de la XOR funcional in vivo. Xantina Oxidasa: constituye el 20 % restante de la XOR funcional normalmente.
Forma inactiva: Desulfo y demolibdo editar
El 60 % de la XOR purificada de leche bovina se encuentra en forma inactiva y existe en 2 formas: Demolibdo: Carece de molibdeno, posiblemente de la molécula de molibdopteria y comúnmente constituye el 40 % de la enzima inactiva. Desulfo: Constituye del 30 % al 40 % restante de la enzima que contiene molibdeno.[2]
Distribución editar
En los mamíferos las mayores concentraciones de estas enzimas se encuentran en el hígado e intestino, sin embargo, hay una variación considerable interespecie que nos indica un amplio rango encontrado en sangre y corazón. Diversos autores usaron técnicas inmunológicas usando anticuerpos anti-XOR, con las cuales se ha detectado XO humana en hepatocitos, sobre todo en región periportal, y las células de Kuffer en el hígado; enterocitos y células globet de yeyuno; glándulas mamarias; células endoteliales de músculo esquelético y riñón; macrófagos y células cebadas; y células epiteliales del tracto biliar, siendo este un indicativo de secreción de XOR hacia la bilis. Estudios de células endoteliales humanas en cultivo, usando anticuerpo anti-XOR de leche humana policlonal fluorescente purificado, mostraron que la enzima se encuentra ubicada tanto en el citosol como en la superficie externa de membrana celular, con predominio en las regiones de contacto intercelular en esta última localización. Se ha sugerido, a partir del uso de anticuerpo monoclonal de ratón en células endoteliales y epiteliales humanas, la localización intravesicular de la XOR intracelularmente, lo cual se ha planteado que pudiera constituir una etapa de almacenamiento previo a su secreción.[2]
Mecanismo catalítico editar
El sitio activo de la xantina oxidasa está compuesto por una unidad de molibdopterina con el átomo de molibdeno también en coordinación con el oxígeno terminal, átomos de azufre y un hidróxido terminal.[3] En la reacción con xantina para formar ácido úrico, un átomo de oxígeno es transferido del molibdeno a la xantina. La reformación del centro activo de molibdeno ocurre por la adición de agua. Al igual que otras oxidoreductasas conteniendo molibdeno, el átomo de oxígeno introducido al substrato por la xantina oxidasa se origina del agua en lugar del dioxígeno (O2).
Importancia clínica editar
En humanos, la xantina oxidasa es encontrada normalmente en el hígado y no libre en la sangre. Durante un daño severo al hígado, la xantina oxidasa es liberada a la sangre, por lo cual un ensayo para xantina oxidasa en sangre es una forma de determinar si ha ocurrido daño hepático.
Debido a que la xantina oxidasa es una ruta metabólica para la formación de ácido úrico, el inhibidor de xantina oxidasa, alopurinol es utilizado en el tratamiento de la gota. Como la xantina oxidasa está involucrada en el metabolismo de la 6-mercaptopurina, se deben tomar recaudos antes de administrar alopurinol en conjunto con 6-mercaptopurina, o su prodroga azatioprina.
La xantinuria es una enfermedad genética poco común, donde la ausencia de xantina oxidasa lleva a una elevada concentración de xantina en sangre y puede provocar problemas de salud tales como una falla renal. No existe un tratamiento específico para este desorden, a los enfermos se le indica evitar los alimentos ricos en purinas y mantener una alta ingesta de líquidos.
Se ha propuesto la inhibición de xantina oxidasa como un mecanismo para mejorar la salud cardiovascular y los síntomas ligados al síndrome de Lesch Nyhan.[4]
Inhibidores editar
Dentro de los inhibidores de la xantina oxidasa se encuentra el alopurinol,[5] oxipurinol,[6] y ácido pítico.[7] Más recientemente, en 2008, se inició la comercialización de un inhibidor de la xantina oxidasa más eficaz en cuanto a reducción del ácido úrico, el febuxostat.
Referencias editar
- ↑ Enroth C, Eger BT, Okamoto K, Nishino T, Nishino T, Pai EF (septiembre de 2000). «Crystal structures of bovine milk xanthine dehydrogenase and xanthine oxidase: structure-based mechanism of conversion». Proc. Natl. Acad. Sci. U.S.A. 97 (20): 10723-8. PMC 27090. PMID 11005854. doi:10.1073/pnas.97.20.10723.
- ↑ a b c d e f g h Mendoza Coussette, Ulises (2005). «Xantina oxidorreductasa, propiedades, funciones y regulación de su expresión genética». Instituto de Ciencias Básicas y Preclínicas “Victoria de Girón”. Rev Cubana Invest Biomed. Consultado el 24 de abril de 2017.
- ↑ Hille R. (2006). «Structure and Function of Xanthine Oxidoreductase». European Journal of Inorganic Chemistry 2006 (10): 1905-2095. doi:10.1002/ejic.200600087.
- ↑ Dawson J, Walters M (octubre de 2006). «Uric acid and xanthine oxidase: future therapeutic targets in the prevention of cardiovascular disease?». British Journal of Clinical Pharmacology 62: 633. PMC 1885190. PMID 17052251. doi:10.1111/j.1365-2125.2006.02785.x.
- ↑ Pacher P, Nivorozhkin A, Szabó C (marzo de 2006). «Therapeutic effects of xanthine oxidase inhibitors: renaissance half a century after the discovery of alopurinol». Pharmacol. Rev. 58 (1): 87-114. PMC 2233605. PMID 16507884. doi:10.1124/pr.58.1.6.
- ↑ Spector T (enero de 1988). «Oxypurinol as an inhibitor of xanthine oxidase-catalyzed production of superoxide radical». Biochem. Pharmacol. 37 (2): 349-52. PMID 2829916. doi:10.1016/0006-2952(88)90739-3.
- ↑ Muraoka S, Miura T (febrero de 2004). «Inhibition of xanthine oxidase by phytic acid and its antioxidative action». Life Sci. 74 (13): 1691-700. PMID 14738912. doi:10.1016/j.lfs.2003.09.040.
 Datos: Q412509
Datos: Q412509