Vida marina
La vida marina, la conforman las plantas, los animales y otros organismos que viven en el agua salada de los mares y océanos, o el agua salobre de los estuarios costeros. En un nivel fundamental, la vida marina ayuda a determinar la naturaleza misma de nuestro planeta. Los organismos marinos producen gran parte del oxígeno que respiramos. Las costas están en parte conformadas y protegidas por la vida marina, y algunos organismos marinos incluso ayudan a crear nuevas tierras.


La mayoría de las formas de vida evolucionaron inicialmente en hábitats marinos. Por volumen, los océanos proporcionan aproximadamente el 90 % de la superficie habitable del planeta.[1] Los primeros vertebrados aparecieron en forma de peces, que viven exclusivamente en agua. Algunos de estos evolucionaron en anfibios que pasan partes de sus vidas en agua y en tierra. Otros peces evolucionaron en mamíferos terrestres y posteriormente regresaron al océano como focas, delfines o ballenas. Las plantas como algas marinas y algas crecen en el agua y son la base de algunos ecosistemas submarinos. El plancton, y particularmente el fitoplancton, son productores primarios claves que forman la base general de la cadena alimentaria oceánica.
Los vertebrados marinos necesitan oxígeno para sobrevivir, y lo obtienen de diversas maneras. Los peces tienen branquias en lugar de pulmones, aunque algunas especies de peces, como el pez pulmonado, tienen ambas. Los mamíferos marinos, tales como delfines, ballenas, nutrias y focas necesitan emerger periódicamente para respirar aire. Algunos anfibios pueden absorber oxígeno a través de su piel. Los invertebrados exhiben una amplia gama de modificaciones para sobrevivir en aguas pobremente oxigenadas, incluyendo tubos de respiración (ver sifones de insectos y moluscos) y branquias (Carcinus). Sin embargo, a medida que la vida de los invertebrados evolucionó en un hábitat acuático, la mayoría tiene poca o ninguna especialización para la respiración en el agua.
En total, hay 230 000 especies marinas documentadas, incluyendo más de 16 000 especies de peces, y se ha estimado que casi dos millones de especies marinas aún no se han documentado.[2] Las especies marinas varían en tamaño desde microscópicas, que incluyen plancton y fitoplancton que pueden ser tan pequeñas con 0,02 micrómetros, hasta grandes cetáceos (ballenas, delfines y marsopas) que en el caso de la ballena azul alcanzan hasta 33 m de longitud, siendo el animal más grande.[3][4]
Agua editar

No hay vida sin agua, que se ha caracterizado como el «disolvente de la vida».[5] El ganador del premio Nobel Albert Szent-Györgyi se refirió al agua como el mater y matrix, la madre y el útero de la vida.[6]
La abundancia de agua en la superficie de la tierra es una característica única que distingue a la Tierra de otros planetas en el sistema solar. La hidrosfera de la Tierra consiste principalmente de los océanos, pero técnicamente incluye todas las superficies de agua en el mundo, incluidos mares interiores, lagos, ríos y aguas subterráneas hasta una profundidad de 2000 m. La ubicación submarina más profunda es el abismo de Challenger de la fosa de las Marianas en el océano Pacífico con una profundidad de 10 911,4 m.[nota 1][7]
La masa de los océanos es de aproximadamente 1.35×1018 toneladas métricas, o aproximadamente 1/4400 de la masa total de la Tierra. Los océanos cubren un área de 3.6×108km² con una profundidad media de 3682 m, lo que resulta en un volumen estimado de 1.3×109km³.[8] Si toda la superficie de la corteza terrestre estuviera a la misma elevación que una esfera lisa, la profundidad del océano mundial resultante sería de 2,7−2,8 km.[9][10]
Alrededor del 97.5 % del agua es salina; el 2.5 % restante es agua dulce. La mayor parte del agua dulce, alrededor del 68,7 %, está presente en forma de hielo en los casquetes de hielo y los glaciares.[11] La salinidad promedio de los océanos de la Tierra es de aproximadamente 35 gramos de sal por kilogramo de agua de mar (3,5 % de sal).[12]
Evolución editar
La edad de la Tierra es unos 4540 millones de años.[13][14][15] La más antigua evidencia concluyente de vida en la Tierra se remonta al menos a hace 3500 millones de años,[16][17] durante la era Eoarcaica luego de que la corteza geológica se comenzara a solidificar luego del eon Hadeano fundido primitivo. Se han encontrado fósiles de tapetes microbianos de 3480 millones de años en areniscas en Australia Occidental.[18][19][20] Otra evidencia física primigenia de una substancia biogénica es el grafito descubierto en rocas metasedimentarias de 3700 millones de años de Groenlandia Occidental[21] como también "restos de material biótico" se han encontrado en rocas de 4100 millones de años de antigüedad en Australia Occidental.[22][23] Uno de los investigadores expresó que «Si la vida se desarrolló con relativa rapidez en la Tierra… entonces puede ser común en el universo».[22]
Todos los organismos en la Tierra descienden de un ancestro común o de un pool de genes ancestral.[24][25] Se cree que reacciones químicas muy energéticas produjeron moléculas auto-replicantes hace unos 4000 millones de años, y unos 500 millones de años después existió el último antepasado común universal.[26] El consenso científico actual es que la bioquímica compleja que hace posible la vida devino de reacciones químicas simples.[27] El comienzo de la vida puede que haya incluido moléculas auto-reproducibles tales como RNA[28] y el desarrollo de células simples.[29]
Las especies actuales son una etapa en el proceso de evolución, su diversidad es el producto de una larga serie de eventos de especialización y extinción.[30] La descendencia común de los organismo fue inicialmente deducida a partir de cuatro verdades simples sobre los organismos: Primero, los mismos poseen distribuciones geográficas que no pueden ser explicadas por procesos de adaptación local. Segundo, la diversidad de la vida no es un conjunto de organismos completamente disjuntos, sino de organismos que comparten similitudes morfológicas. Tercero, rasgos vestigiales sin un propósito claro se asemejan a los rasgos ancestrales funcionales y, finalmente, los organismos se pueden clasificar utilizando estas similitudes en una jerarquía de grupos anidados, similar a un árbol genealógico.[31] Sin embargo, la investigación moderna ha sugerido que, debido a la transferencia horizontal de genes, este «árbol de la vida» puede ser más complicado que un árbol de ramificación simple ya que algunos genes se han diseminado independientemente entre especies distantes relacionadas.[32][33] Las especies del pasado también han dejado registros de su historia evolutiva. Los fósiles, junto con la anatomía comparada de los organismos actuales, constituyen el registro morfológico o anatómico.[34] Al comparar las anatomías de las especies modernas y extintas, los paleontólogos pueden inferir los linajes de esas especies. Sin embargo, este enfoque es más exitoso para los organismos que tienen partes duras del cuerpo, tales como conchas, huesos o dientes. Además, debido a que los procariotas, tales como las bacterias y las arqueas, comparten un conjunto limitado de morfologías comunes, sus fósiles no proporcionan información sobre sus ancestros.

Más recientemente, la evidencia de descendencia común proviene del estudio de similitudes bioquímicas entre organismos. Por ejemplo, todas las células vivas usan el mismo conjunto básico de nucleótidos y aminoácidos.[35] El desarrollo de la genética molecular ha revelado el registro de la evolución que queda en los genomas de los organismos: permitiendo realizar una datación de cuando las especies divergieron mediante el reloj molecular producido por las mutaciones. .[36] Por ejemplo, estas comparaciones de secuencias de ADN han revelado que los humanos y los chimpancés comparten el 98% de sus genomas y el analizar las pocas áreas donde difieren ayuda a determinar sobre cuándo existió el ancestro común de estas especies.[37]
Los procariotas habitaron la Tierra hace aproximadamente 3 a 4000 millones de años.[38][39] No se produjeron cambios obvios en la morfología o la organización celular en estos organismos durante los siguientes pocos miles de millones de años.[40] Las células eucariotas surgieron hace unos 1,6 a 2,7 mil millones de años. El siguiente gran cambio en la estructura celular se produjo cuando las bacterias fueron engullidas por células eucarióticas, en una asociación cooperativa llamada endosimbiosis.[41][42] Las bacterias capturadas y la célula huésped luego experimentaron una coevolución, con la bacteria evolucionando para generar mitocondrias o hidrogenosomas.[43] Otra conjunción de organismos de tipo cianobacteriano condujo a la formación de cloroplastos en algas y plantas. .[44]
La historia de la vida fue la de los eucariotes, procariotas y arqueas unicelulares, hasta hace unos 610 millones de años cuando comenzaron a aparecer organismos multicelulares en los océanos en el período de Ediacara.[38][45] La evolución de la multicelularidad se produjo en múltiples eventos independientes, en organismos tan diversos como esponjas, algas marrones, cianobacterias, mohos de limo y mixobacterias.[46] En enero de 2016, los científicos informaron que, hace unos 800 millones de años, un cambio genético menor en una sola molécula llamada GK-PID puede haber permitido que los organismos pasen de un único organismo celular a uno constituido por muchas células.[47] Poco después de la aparición de estos primeros organismos multicelulares, apareció una notable cantidad de diversidad biológica durante aproximadamente 10 millones de años, en un evento denominado explosión cámbrica. En ella, la mayoría de los tipos de animales modernos aparecen en el registro fósil, así como linajes únicos que posteriormente se extinguieron.[48] Se han propuesto varios desencadenantes para la explosión del Cámbrico, incluida la acumulación de oxígeno en la atmósfera a partir de la fotosíntesis.[49]
Hace unos 500 millones de años, las plantas y los fungi comenzaron a colonizar la tierra. La evidencia de la aparición de las primeras plantas terrestres ocurre en el Ordovícico, hace alrededor de 450 millones de años, en forma de esporas fósiles.[50] Las plantas terrestres comenzaron a diversificarse a fines del Silúrico, hace alrededor de 430 millones de años.[51] La colonización de la tierra por las plantas pronto fue seguida por artrópodos y otros animales.[52] Los insectos fueron particularmente exitosos e incluso hoy en día constituyen la mayoría de las especies animales[53] Los anfibios aparecieron por primera vez hace 364 millones de años, seguidos por amniotes tempranos y aves hace alrededor de 155 millones de años (ambos de linajes similares a "reptiles"), mamíferos hace 129 millones de años, homínidos hace unos 10 millones de años y humanos modernos hace unos 250 000 años.[54][55][56] Sin embargo, a pesar de la evolución de estos animales grandes, los organismos más pequeños similares a los tipos que evolucionaron temprano en este proceso continúan teniendo mucho éxito y dominan la Tierra, con la mayoría de la biomasa y de las especie siendo procariotas.[57]
Las estimaciones sobre el número de especies actuales de la Tierra varían de 10 a 14 millones,[58] de los cuales se han documentado alrededor de 1,2 millones y más del 86 por ciento aún no se han descrito.[59]
Microorganismos editar


Los microorganismos representan aproximadamente el 70% de la biomasa marina.[60] Un microorganismo, o microbio, es un organismo microscópico tan pequeño que no puede ser detectado a simple vista. Puede ser unicelular[61] o multicelular. Los microorganismos son diversos e incluyen todas las bacterias y archaea, la mayoría de los protozoos tales como algas, hongos y ciertos animales microscópicos tales como rotíferos.
Muchos animales microscópicos y plantas tienen etapas juveniles microscópicas. Algunos microbiólogos también clasifican a los virus (y viroides) como microorganismos, aunque otros consideran que estos no son seres vivos.[62][63]
Los microorganismos son cruciales para el reciclaje de nutrientes en los ecosistemas, ya que actúan como descomponedores. Algunos microorganismos son patógenos y causan enfermedades e incluso la muerte en plantas y animales.[64] Como habitantes del medio ambiente más grande de la Tierra, los sistemas marinos microbianos impulsan cambios en todos los sistemas globales. Los microbios son responsables de prácticamente toda la fotosíntesis que ocurre en el océano, así como del ciclo del carbono, nitrógeno, fósforo, otros nutrientes y oligoelementos.[65]

| Microorganismos marinos |
| ||||||||||||||||||||||||
_-_Stephanopyxis_sp._-_630x_(14117161428).jpg)

La vida microscópica submarina es diversa y aún no se comprende bien, como por ejemplo en lo que respecta al papel de los virus en los ecosistemas marinos.[66] La mayoría de los virus marinos son bacteriófagos, que son inofensivos para las plantas y los animales, pero son esenciales para la regulación de los ecosistemas de agua dulce y salada.[67] Infectan y destruyen bacterias en comunidades microbianas acuáticas y son el mecanismo más importante de reciclaje de carbono en el medio marino. Las moléculas orgánicas liberadas por las células bacterianas muertas estimulan el crecimiento de algas y bacterias frescas.[68] La actividad viral también puede contribuir a la bomba biológica, el proceso por el cual el carbono es secuestrado en las profundidades del océano.[69]


Una corriente de microorganismos transportados por el aire rodea el planeta por encima de los sistemas meteorológicos pero por debajo de las rutas aéreas comerciales.[70] Algunos microorganismos peripatéticos son arrastrados por las tormentas de polvo terrestres, pero la mayoría se origina a partir de microorganismos marinos en el rocío del mar. En 2018, los científicos informaron que cientos de millones de virus y decenas de millones de bacterias se depositan diariamente en cada metro cuadrado del planeta.[71][72]
Los organismos microscópicos viven en toda la biosfera. La masa de microorganismos procariotas, que incluye bacterias y arqueas, pero no los microorganismos eucariotas nucleados, puede llegar a 800 millones de toneladas de carbono (de la masa total de la biosfera, estimada entre 1000 y 4000 millones de toneladas).[73] Se han encontrado microbios marinos barófilos unicelulares a una profundidad de 10.900 m en la Fosa de las Marianas, el lugar más profundo de los océanos de la Tierra.[74][75] Los microorganismos viven dentro de las rocas a 580 m por debajo del lecho marino bajo 2590 m de océano frente a la costa del noroeste de los Estados Unidos,[74][76] así como a 2400 m debajo del lecho marino frente a Japón.[77] La temperatura más alta conocida a la que puede existir vida microbiana es 122 °C (Methanopyrus kandleri).[78] En 2014, los científicos confirmaron la existencia de microorganismos que vivían a 800 m por debajo del hielo de la Antártida.[79][80] Según un investigador, "puedes encontrar microbios en todas partes; son extremadamente adaptables a las condiciones y sobreviven donde sea que estén".[74]
Protistas marinos editar
Los protistas son eukaryotas que no pueden ser clasificados como plantas, hongos o animales. Suelen ser unicelulares y microscópicos. La vida se originó como procariotas unicelulares (bacterias y arqueas) y luego evolucionó en eucariotas más complejos. Los eucariotas son las formas de vida más desarrolladas conocidas que comprende plantas, animales, hongos y protistas. El término protista se utilizó históricamente como un término de conveniencia para eucariotas que no pueden clasificarse estrictamente como plantas, animales u hongos. No son parte de la cladística moderna, porque son parafiléticos (que carecen de un ancestro común). Los protistas se pueden dividir ampliamente en cuatro grupos dependiendo de si su nutrición es similar a la de una planta, un animal o un hongo,[81] o una mezcla de ellas.[82]
Protistas de acuerdo a cómo se procuran su alimento
| |||||||
|---|---|---|---|---|---|---|---|
| Tipo de protista | Descripción | Ejemplo | Otros ejemplos | ||||
| Tipo planta | Los protistas autotróficos producen su propio alimento sin necesidad de consumir otros organismos, por lo general realizando fotosíntesis | 
|
Alga roja, Cyanidium sp. | Alga verde, alga marrón, diatomeas y algunas dinoflagelados. Los protistas tipo plantas son componentes importantes del fitoplancton. | |||
| Tipo animal | Protistas heterotróficos que obtienen su alimento consumiendo otros organismos | 
|
Protista radiolario dibujado por Haeckel | Foraminiferos, y algunas amebas, ciliados y flagelados marinos. | |||
| Tipo hongo | Protistas saprotróficos que obtienen su alimento de los restos de organismos que se han roto y decaído. | 
|
Los laberintúlidos marinos forman redes laberínticas de tubos por los que pueden desplazarse las amebas sin pseudópodos. | Liquen marino | |||
| Mixotrofos | Varios
|
Protistas mixotróficos y osmotróficos que obtienen su alimento de una combinación de las anteriores | _(cropped).jpg)
|
Euglena mutabilis un flagelado fotosintético | Existen muchos mixótropos entre los protistas, incluidos entre los ciliates, rhizaria y dinoflagelados[83] | ||


Los protistas son organismos muy diversos actualmente organizados en 18 filos, pero no resulta fácil clasificarlos.[85][86] Los estudios muestran que existe una alta diversidad de protistas en los océanos, los respiraderos de aguas profundas y los sedimentos de los ríos, lo que sugiere que aún no se ha descubierto una gran cantidad de comunidades microbianas eucariotas.[87][88] Ha habido poca investigación sobre protistas mixotróficos, pero estudios recientes en ambientes marinos encontraron que los protistas mixotróficos contribuyen con una parte significativa de la biomasa protista.[83]
- Protistas unicelulares y microscópicos
-
![Las diatomeas son un gran grupo de algas que genera el 20% de la producción mundial de oxígeno.[89]](//upload.wikimedia.org/wikipedia/commons/thumb/3/31/Diatoms_through_the_microscope.jpg/297px-Diatoms_through_the_microscope.jpg)
-
![Las diatomeas tienen paredes celulares similares al vidrio hechas de sílice y se llaman frústulas.[90]](//upload.wikimedia.org/wikipedia/commons/thumb/9/90/Diatom_algae_Amphora_sp.jpg/260px-Diatom_algae_Amphora_sp.jpg)
-
 Frústula en un fósil de diatomea de hace 32 a 40 millones de años
Frústula en un fósil de diatomea de hace 32 a 40 millones de años -

-
 Alga monocelular, Gephyrocapsa oceanica
Alga monocelular, Gephyrocapsa oceanica -
 Dos dinoflagelados
Dos dinoflagelados -
 Zooxanthellae es un alga fotosintética que vive dentro de receptores como el coral.
Zooxanthellae es un alga fotosintética que vive dentro de receptores como el coral. -
 Un ciliado unicelular viviendo dentro de una Zoochlorella verde en forma endosimbiótica.
Un ciliado unicelular viviendo dentro de una Zoochlorella verde en forma endosimbiótica. -

-
 Este ciliado digiere una cianobacterias. El citostoma o boca se encuentra abajo a la derecha.
Este ciliado digiere una cianobacterias. El citostoma o boca se encuentra abajo a la derecha.
![Las diatomeas son un gran grupo de algas que genera el 20% de la producción mundial de oxígeno.[89]](/wiki/Archivo:Diatoms_through_the_microscope.jpg)
![Las diatomeas tienen paredes celulares similares al vidrio hechas de sílice y se llaman frústulas.[90]](/wiki/Archivo:Diatom_algae_Amphora_sp.jpg)

.jpg)
.jpg)





Microanimales marinos editar
Como juveniles, los animales se desarrollan a partir de estadios microscópicos, que pueden incluir esporas, huevos y larvas. Al menos un grupo de animales microscópicos, el parásito cnidario Myxozoa, es unicelular en su forma adulta e incluye especies marinas. Otros microanimales marinos adultos son multicelulares. Los artrópodos adultos microscópicos se encuentran más comúnmente tierra adentro en agua dulce, pero también hay especies marinas. Los crustáceos marinos adultos microscópicos incluyen algunos copépodos, cladóceros y tardígrados (osos de agua). Algunos nematodos y rotíferos marinos también son demasiado pequeños para ser reconocidos a simple vista, al igual que muchos loricifera, incluida las especies recientemente descubiertas de organismos anaeróbicos que pasan sus vidas en un ambiente de anóxico.[91][92] Los copépodos contribuyen más a la productividad secundaria y al sumidero de carbono de los océanos del mundo que cualquier otro grupo de organismos.
- Microanimales marinos
-
 Más de 10,000 especies marinas son copépodos, pequeños crustáceos a menudo micróscopicos
Más de 10,000 especies marinas son copépodos, pequeños crustáceos a menudo micróscopicos -
 Fotografía en campo oscuro de un gastrotrico, un animal tipo lombriz que vive entre las partículas sedimentarias
Fotografía en campo oscuro de un gastrotrico, un animal tipo lombriz que vive entre las partículas sedimentarias -
 Pliciloricus enigmaticus blindado, de 0.2 mm de largo, vive en espacios entre la arena marina.
Pliciloricus enigmaticus blindado, de 0.2 mm de largo, vive en espacios entre la arena marina. -
 Dibujo de un tardígrado (oso de agua) en un grano de arena
Dibujo de un tardígrado (oso de agua) en un grano de arena -





_-_160x_(13402418244).jpg)
Algas y plantas marinas editar

Las algas y las plantas microscópicas proporcionan hábitats importantes para la vida, a veces actuando como lugares de escondite y búsqueda de alimento para las formas larvarias de peces e invertebrados más grandes. Las algas se encuentran muy difundidas en los océanos y adoptan formas diversas. Las algas fotosintéticas microscópicas contribuyen con una mayor proporción de la producción fotosintética del mundo que la de todos los bosques terrestres combinados. La mayor parte del nicho ocupado por subplantas en la tierra en realidad está ocupado por algas macroscópicas en el océano, tales como Sargassum y algas marinas, que crean bosques de algas marinas.
Las plantas que sobreviven en el mar a menudo lo hacen en aguas poco profundas, tales como los pastosmarinos (ejemplos de los cuales son zacate, Zostera, y el pasto tortuga, Thalassia). Estas plantas se han adaptado a los elevados niveles de salinidad del medio ambiente oceánico. La zona intermareal también es un buen sitio para el desarrollo de la vida en el mar, donde pueden crecer manglares o spartina o ammofila.
-

-
 Volvox es un alga verde microscópica de agua dulce con simetría esférica. Se puede observar colonias jóvenes desarrollándose dentro de otras más grandes.
Volvox es un alga verde microscópica de agua dulce con simetría esférica. Se puede observar colonias jóvenes desarrollándose dentro de otras más grandes. -

-
 Bosque de algas marinas
Bosque de algas marinas




Invertebrados editar



Los primeros animales fueron invertebrados marinos, es decir, los vertebrados llegaron más tarde. Los animales son eucariotas multicelulares, y se distinguen de las plantas, las algas y los hongos por la falta de paredes celulares. Los invertebrados marinos son animales que habitan en un ambiente marino aparte de los miembros vertebrados del filo cordado. Los invertebrados carecen de columna vertebral. Algunos han evolucionado una concha o un exoesqueleto duro.
Los primeros fósiles de animales probablemente pertenezcan al género Dickinsonia, y se remontan a hace unos 571 millones a 541 millones de años. La Dickinsonia individual se asemeja a un óvalo acanalado bilateralmente simétrico. Continuaron creciendo hasta que se cubrieron con sedimentos o se extinguieron de otra manera, y pasaron la mayor parte de sus vidas con sus cuerpos firmemente anclados al sedimento. Sus afinidades taxonómicas son actualmente desconocidas, pero su modo de crecimiento es consistente con una afinidad bilateral.
Aparte de la Dickinsonia, otros fósiles de animales ampliamente aceptados como muy antiguos son los cnidarios de apariencia más bien moderna (el grupo que incluye medusas, anémonas de mar e hidra), posiblemente de hace unos 580 millones de años. La biota de Ediacara, que floreció durante los últimos 40 millones de años antes del inicio del Cámbrico, fueron los primeros animales de más de unos pocos centímetros de largo. Como Dickinsonia, muchos eran planos con una apariencia "acolchada" y parecían tan extraños que hubo una propuesta para clasificarlos como un reino separado, Vendozoa. Otros, sin embargo, han sido interpretados como moluscos tempranos (Kimberella), equinodermos (Arkarua) y artrópodos (Spriggina, Parvancorina). Todavía se debate sobre la clasificación de estos especímenes, principalmente porque las características de diagnóstico que permiten a los taxónomos clasificar organismos más recientes, como las similitudes con organismos vivos, generalmente están ausentes en los ediacaranos. Sin embargo, parece haber pocas dudas de que Kimberella era al menos un animal bilateriano triploblástico, en otras palabras, un animal significativamente más complejo que los cnidarios.
La fauna de conchas pequeñas es un conjunto muy variado de fósiles que se encuentran entre los períodos Ediacarano tardío y Cámbrico Medio. La más temprana, Cloudina, presenta signos de defensa exitosa contra la depredación y puede indicar el comienzo de una carrera armamentista evolutiva. Algunas pequeñas conchas del Cámbrico temprano casi seguramente pertenecían a los moluscos, mientras que los dueños de algunas "placas de armadura", Halkieria y Microdictyon, fueron finalmente identificados cuando se encontraron especímenes más completos en lagertätten del Cámbrico que conservaban animales de cuerpo blando.
En la década de 1970 ya existía un debate acerca de si el surgimiento de los filos modernos fue "explosivo" o gradual, pero oculto por la escasez de fósiles de animales precámbricos. Un nuevo análisis de los fósiles de Burgess Shale lagerstätte aumentó el interés por el tema cuando reveló animales, como Opabinia, que no encajaba en ningún phylum conocido. En aquel momento e interpretaron como evidencia de que los filos modernos habían evolucionado muy rápidamente en la explosión del Cámbrico y que las "extrañas maravillas" de Burgess Shale mostraban que el Cámbrico Temprano fue un período singularmente experimental de evolución animal. Los descubrimientos posteriores de animales similares y el desarrollo de nuevos enfoques teóricos llevaron a la conclusión de que muchas de las "maravillas extrañas" eran "tías" evolutivas o "primos" de grupos modernos, por ejemplo, que Opabinia era miembro de los lobópodos. Un grupo que incluye a los antepasados de los artrópodos, y que puede haber estado estrechamente relacionado con los tartigrados modernos. Sin embargo, todavía hay mucho debate sobre si la explosión del Cámbrico fue realmente explosiva y, en caso afirmativo, cómo y por qué ocurrió y por qué parece única en la historia de los animales.
Los invertebrados se agrupan en diferentes filos. Informalmente se puede pensar en los filos como una forma de agrupar organismos de acuerdo con su plan corporal. Un plan corporal se refiere a un plano que describe la forma o morfología de un organismo, como por ejemplo su simetría, segmentación y la disposición de sus apéndices. La idea de los planes corporales se originó con los vertebrados, que se agruparon en un solo filo. Pero el plan del cuerpo de los vertebrados es solo uno de muchos, y en los invertebrados existen muchos planes de cuerpo o de filos diferentes. La historia del descubrimiento de los planes corporales puede verse como un movimiento desde una cosmovisión centrada en los vertebrados, hasta ver a los vertebrados como un plan corporal entre muchos. Entre los zoólogos pioneros, Linneo identificó dos planes corporales fuera de los vertebrados; Cuvier identificó tres; y Haeckel tuvo cuatro, así como el Protista con ocho más, para un total de doce. A modo de comparación, el número de filos reconocidos por los zoólogos modernos ha aumentado a 35.
Históricamente, se pensó que los planes corporales habían evolucionado rápidamente durante la explosión del Cámbrico, pero una comprensión más amplia de la evolución animal sugiere un desarrollo gradual de los planes corporales a lo largo del Paleozoico temprano y más allá. De manera más general, un filo se puede definir de dos maneras: como se describió anteriormente, como un grupo de organismos con un cierto grado de similitud morfológica o de desarrollo (la definición fenética), o un grupo de organismos con un cierto grado de relación evolutiva (la definición filogenética).
Los artrópodos suman aproximadamente 1 113 000 especies descritas, los moluscos alrededor de 85 000 y los cordados alrededor de 52 000.
Esponjas marinas editar
.jpg)
Las esponjas son animales del filo Porifera (latín moderno: que posee poros). Son organismos multicelulares que tienen cuerpos llenos de poros y canales que permiten que el agua circule a través de ellos. Su cuerpo consiste de mesoílo gelatinoso intercalado entre dos capas delgadas de células. Tienen células no especializadas que pueden transformarse en otros tipos y que a menudo migran entre las capas celulares principales y el mesoil en el proceso. Las esponjas no tienen sistemas nervioso, digestivo o circulatorio. En cambio, la mayoría mantiene un flujo constante de agua a través de sus cuerpos para obtener alimentos y oxígeno y para eliminar los desechos.
Las esponjas son similares a otros animales porque son multicelulares, heterótrofas, carecen de paredes celulares y producen células de esperma. A diferencia de otros animales, carecen de tejidos y órganos verdaderos, y no tienen simetría corporal. Las formas de sus cuerpos están adaptadas para la máxima eficiencia del flujo de agua a través de la cavidad central, donde deposita los nutrientes, y sale a través de un agujero llamado osculum. Muchas esponjas tienen esqueletos internos de esponjas y/o espículas de carbonato de calcio o dióxido de silicio. Todas las esponjas son animales acuáticos sésiles. Aunque hay especies de agua dulce, la gran mayoría son especies marinas (agua salada), que van desde zonas de mareas hasta profundidades que superan los 8800 m.
Si bien la mayoría de las aproximadamente 5000−10 000 especies conocidas de esponjas se alimentan de bacterias y otras partículas de alimentos en el agua, algunas albergan microorganismos de fotosíntesis tales como endosimbiontes y estas alianzas a menudo producen más alimentos y oxígeno de los que consumen. Algunas especies de esponjas que viven en ambientes pobres en alimentos se han convertido en carnívoras que se alimentan principalmente de pequeños crustáceos.
-
 Biodiversidad de las esponjas. En la fotografía se observan cuatro especies de esponjas.
Biodiversidad de las esponjas. En la fotografía se observan cuatro especies de esponjas. -

-
 Canasta de flores de Venus a una profundidad de 2572 m
Canasta de flores de Venus a una profundidad de 2572 m -

-


.jpg)
.jpg)
.jpg)
.jpg)
Linneo identificó erróneamente las esponjas como plantas en el orden Algas. Durante mucho tiempo después, las esponjas fueron asignadas a un subreino separado, Parazoa (que significa al lado de los animales). Ahora están clasificadas como un filo parafilético a partir del cual los animales superiores han evolucionado.
Cnidarios marinos editar

Los cnidarios (en griego ortiga) se distinguen por la presencia de células urticantes, células especializadas que utilizan principalmente para capturar presas. Los cnidarios incluyen corales, anémonas de mar, medusas e hidrozoos. Forman un filo que contiene más de 10 000 especies de animales que se encuentran exclusivamente en ambientes acuáticos (principalmente marinos). Sus cuerpos consisten de mesoglea, una sustancia gelatinosa no viva, intercalada entre dos capas de epitelio que son en su mayoría de una célula de espesor. Tienen dos formas corporales básicas: medusas natatorias y pólipos sésiles, ambos son radialmente simétricos con bocas rodeadas de tentáculos que contienen cnidocitos. Ambas formas tienen un único orificio y una cavidad corporal que se utilizan para la digestión y la respiración.
Se han encontrado cnidarios fósiles en rocas formadas hace unos 580 millones de años. Los fósiles de cnidarios que no construyen estructuras mineralizadas son raros. Los científicos actualmente piensan que los cnidarios, ctenóforos y bilaterales están más estrechamente relacionados con las esponjas calcáreas que estas con otras esponjas, y que los antozoos son las "tías" o "hermanas" evolutivas de otros cnidarios, y están más estrechamente relacionados con los bilaterales.
Los cnidarios son los animales más simples en los que las células se organizan en tejidos. La anémona de mar estrellada se utiliza como organismo modelo en la investigación. Es fácil de cuidar en el laboratorio y se ha desarrollado un protocolo que puede producir una gran cantidad de embriones a diario. Existe un notable grado de similitud en la conservación de secuencias genéticas y la complejidad entre la anémona de mar y los vertebrados. En particular, los genes asociados a la formación de la cabeza en vertebrados también están presentes en la anémona.
-
 Las anémonas marinas son comunes en los pozas de marea
Las anémonas marinas son comunes en los pozas de marea -
 Sus tentáculos pinchan y pueden paralizar a peces pequeños
Sus tentáculos pinchan y pueden paralizar a peces pequeños -

-
 En la medida que las islas se hunden en el mar, los corales crecen para compensar el nivel en ascenso del agua en el atolón
En la medida que las islas se hunden en el mar, los corales crecen para compensar el nivel en ascenso del agua en el atolón




-

-

-
![Turritopsis dohrnii, una pequeña medusa biológicamente inmortal transfiere sus células a sus descendientes.[94][95]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b0/Turritopsis_dohrnii_%28cropped%29.jpg/180px-Turritopsis_dohrnii_%28cropped%29.jpg) Turritopsis dohrnii, una pequeña medusa biológicamente inmortal transfiere sus células a sus descendientes.[94][95]
Turritopsis dohrnii, una pequeña medusa biológicamente inmortal transfiere sus células a sus descendientes.[94][95] -
![Medusa luna, habita en las aguas costeras en diversas partes del mundo[96]](//upload.wikimedia.org/wikipedia/commons/thumb/c/c4/Aurelia_aurita_%28Cnidaria%29_Luc_Viatour_%28cropped%29.jpg/158px-Aurelia_aurita_%28Cnidaria%29_Luc_Viatour_%28cropped%29.jpg) Medusa luna, habita en las aguas costeras en diversas partes del mundo[96]
Medusa luna, habita en las aguas costeras en diversas partes del mundo[96] -
![Medusa melena de león, la medusa conocida de mayor tamaño[97]](//upload.wikimedia.org/wikipedia/commons/thumb/2/22/Largelionsmanejellyfish.jpg/135px-Largelionsmanejellyfish.jpg) Medusa melena de león, la medusa conocida de mayor tamaño[97]
Medusa melena de león, la medusa conocida de mayor tamaño[97]
.jpg)

![Turritopsis dohrnii, una pequeña medusa biológicamente inmortal transfiere sus células a sus descendientes.[94][95]](/wiki/Archivo:Turritopsis_dohrnii_(cropped).jpg)
![Medusa luna, habita en las aguas costeras en diversas partes del mundo[96]](/wiki/Archivo:Aurelia_aurita_(Cnidaria)_Luc_Viatour_(cropped).jpg)
![Medusa melena de león, la medusa conocida de mayor tamaño[97]](/wiki/Archivo:Largelionsmanejellyfish.jpg)
Equinodermos editar

Los equinodermos (griego: piel espinosa) es un phylum que contiene solo invertebrados marinos. Los adultos son reconocibles por su simetría radial (generalmente de cinco puntos) e incluyen estrellas de mar, erizos de mar, dólares de la arena y pepinos de mar, así como lirios de mar. Los equinodermos se encuentran en un amplio espectro de profundidades de los océanos, desde la zona intermareal a la zona abisal. El phylum contiene alrededor de 7000 especies vivas, por lo que es el segundo grupo más grande de deuterostomes (un superphylum), después de los cordados (que incluyen los vertebrados, como aves, peces, mamíferos y reptiles).
Los equinodermos son únicos entre los animales por tener simetría bilateral en la etapa larvaria, pero cinco simetrías (pentamerismo, un tipo especial de simetría radial) como adultos.
Los equinodermos son importantes tanto biológica como geológicamente. Biológicamente, hay pocos grupos tan abundantes en el desierto biótico de las profundidades marinas, así como en los océanos menos profundos. La mayoría de los equinodermos son capaces de regenerar tejidos, órganos, extremidades y reproducirse asexualmente; en algunos casos, pueden experimentar una regeneración completa de una sola extremidad. Geológicamente, el valor de los equinodermos se encuentra en sus esqueletos osificados, que son importantes contribuyentes a muchas formaciones de piedra caliza y pueden proporcionar pistas valiosas sobre el entorno geológico. Fueron las especies más utilizadas en la investigación regenerativa en los siglos XIX y XX. Además, algunos científicos sostienen que la radiación de los equinodermos fue responsable de la revolución marina mesozoica.
-
 Equinodermo significa literalmente "piel pinchuda", tal como ilustra este erizo de mar melón
Equinodermo significa literalmente "piel pinchuda", tal como ilustra este erizo de mar melón -
![La estrella de mar ocre fue el primer depredador clave en ser estudiado. Limitan las poblaciones de mejillones que pueden abrumar a las comunidades intermareales.[98]](//upload.wikimedia.org/wikipedia/commons/thumb/8/89/Ochre_sea_star.jpg/240px-Ochre_sea_star.jpg) La estrella de mar ocre fue el primer depredador clave en ser estudiado. Limitan las poblaciones de mejillones que pueden abrumar a las comunidades intermareales.[98]
La estrella de mar ocre fue el primer depredador clave en ser estudiado. Limitan las poblaciones de mejillones que pueden abrumar a las comunidades intermareales.[98] -
 Coloridos lirios de mar en aguas poco profundas
Coloridos lirios de mar en aguas poco profundas

![La estrella de mar ocre fue el primer depredador clave en ser estudiado. Limitan las poblaciones de mejillones que pueden abrumar a las comunidades intermareales.[98]](/wiki/Archivo:Ochre_sea_star.jpg)

-
 Los pepinos de mar se alimentan filtrando plancton y sólidos en suspensión en el agua de mar
Los pepinos de mar se alimentan filtrando plancton y sólidos en suspensión en el agua de mar -
 El pepino de mar bentopelágico puede despegar desde el fondo del mar y ascender hasta 1000 m
El pepino de mar bentopelágico puede despegar desde el fondo del mar y ascender hasta 1000 m -
 Este pepino de mar de agua profunda, un cerdo de mar, es el único equinodermo que se desplaza mediante locomoción con patas.
Este pepino de mar de agua profunda, un cerdo de mar, es el único equinodermo que se desplaza mediante locomoción con patas.



Moluscos marinos editar

Los moluscos (latín: suave) forman un phylum con aproximadamente 85 000 especies reconocidas existentes. Son el mayor phylum marino en términos de número de especies, que comprende aproximadamente el 23 % de todos los organismos marinos mencionados. Los moluscos tienen formas más variadas que otros phylums de invertebrados. Son muy diversos, no solo en tamaño y en estructura anatómica, sino también en comportamiento y en hábitat. La mayoría de las especies todavía viven en los océanos, desde las costas hasta la zona abisal, pero algunas forman una parte significativa de la fauna de agua dulce y los ecosistemas terrestres.
El filo de los moluscos está dividido en 9 o 10 clases taxonómicas, dos de las cuales están extintas. Estas clases incluyen gastrópodos, bivalvos y cefalópodos, así como otras clases menos conocidas pero distintivas. Los gastrópodos con conchas protectoras son los denominados caracoles, mientras que los gastrópodos sin conchas protectoras son denominados babosas. Los gastrópodos son, con mucho, los moluscos más numerosos en términos de especies clasificadas, que representan el 80 % del total. Los bivalvos incluyen almejas, ostras, berberechos, mejillones, vieiras y muchas otras familias. Hay alrededor de 8000 especies de bivalvos marinos (incluyendo especies de agua salobre y estuarina), y alrededor de 1200 especies de agua dulce. Los cefalópodos incluyen pulpo, calamar y sepia. Se encuentran en todos los océanos y neurológicamente son los invertebrados más avanzados. Se identificaron alrededor de 800 especies vivas de cefalópodos marinos, y se ha descrito un estimado de 11 000 taxones extintos. No hay cefalópodos exclusivamente de agua dulce.
-
![El calamar colosal, es el más grande de todos los invertebrados[99]](//upload.wikimedia.org/wikipedia/commons/thumb/3/37/Calmarcolossal.jpg/126px-Calmarcolossal.jpg) El calamar colosal, es el más grande de todos los invertebrados[99]
El calamar colosal, es el más grande de todos los invertebrados[99] -
![El nautilus es un fósil viviente que ha cambiado muy poco su aspecto desde que surgiera hace unos 500 millones de años como uno de los primeros cefalópodos.[100][101][102]](//upload.wikimedia.org/wikipedia/commons/thumb/d/d5/Nautilus_Palau.JPG/240px-Nautilus_Palau.JPG) El nautilus es un fósil viviente que ha cambiado muy poco su aspecto desde que surgiera hace unos 500 millones de años como uno de los primeros cefalópodos.[100][101][102]
El nautilus es un fósil viviente que ha cambiado muy poco su aspecto desde que surgiera hace unos 500 millones de años como uno de los primeros cefalópodos.[100][101][102] -
 Los gastrópodos marinos son caracoles marinos o babosas marinas. Este nudibranquio es una babosa marina.
Los gastrópodos marinos son caracoles marinos o babosas marinas. Este nudibranquio es una babosa marina. -
 El caracol marino Syrinx aruanus tiene la mayor concha de todos los gastrópodos en existencia
El caracol marino Syrinx aruanus tiene la mayor concha de todos los gastrópodos en existencia -

-
 El mejillón común, otro bivalvo
El mejillón común, otro bivalvo
![El calamar colosal, es el más grande de todos los invertebrados[99]](/wiki/Archivo:Calmarcolossal.jpg)
![El nautilus es un fósil viviente que ha cambiado muy poco su aspecto desde que surgiera hace unos 500 millones de años como uno de los primeros cefalópodos.[100][101][102]](/wiki/Archivo:Nautilus_Palau.JPG)





Los moluscos tienen formas tan diversas que muchos libros de texto basan sus descripciones de la anatomía de los moluscos en un molusco ancestral generalizado o hipotético. Este molusco generalizado no está segmentado y tiene simetría bilateral con un lado inferior que consiste en un solo pie muscular.[103][104] Adicionalmente, tiene otras tres características clave. En primer lugar, tiene una capa muscular llamada manto que cubre sus vísceras y que contiene una cavidad utilizada para la respiración y la excreción. Una concha secretada por el manto cubre la superficie superior.[104] En segundo lugar (aparte de los bivalvos) tiene una lengua áspera llamada rádula utilizada para alimentarse. En tercer lugar, tiene un sistema nervioso que incluye un complejo sistema digestivo que utiliza pelos microscópicos que funcionan con los músculos, llamados cilios, para exudar la mucosidad. El molusco generalizado tiene dos cuerdas nerviosas pareadas (tres en bivalvos). El cerebro, en las especies que lo tienen, rodea el esófago. La mayoría de los moluscos tienen ojos y todos tienen sensores que detectan químicos, vibraciones y tacto. El tipo más simple de sistema reproductivo de los moluscos se basa en la fertilización externa, pero ocurren variaciones más complejas. Todos producen huevos, de los cuales pueden surgir larvas trocóforas, larvas veliger más complejas o adultos en miniatura. La representación es bastante similar a la de los monoplacóforos modernos, y algunos estudiosos sugieren que puede parecerse a la de los moluscos primigenios.[103]: 284–291 [103]: 298–300 [105][106]
Existe evidencia concluyente sobre la apariencia de los gastrópodos marinos, cefalópodos y bivalvos en el periodo Cámbrico 538,8 a 485,4 millones de años. Sin embargo, la historia evolutiva tanto de la emergencia de los moluscos a partir de los Lophotrochozoa ancestrales y su diversificación hacia las formas existentes en la actualidad y formas fósiles son aun temas de debate entre los científicos.
Artrópodos marinos editar

Los artrópodos (en griego: patas articuladas) tienen un exoesqueleto (esqueleto externo), un cuerpo segmentado y apéndices articulados (apéndices pareados). Forman un phylum que incluye insectos, arácnidos, miriópodos y crustáceos. Los artrópodos se caracterizan por sus extremidades articuladas y su cutícula de quitina, a menudo mineralizada con carbonato de calcio. El plan del cuerpo del artrópodo consiste en segmentos, cada uno con un par de apéndices. La cutícula rígida inhibe el crecimiento, por lo que los artrópodos la reemplazan periódicamente mediante la muda. Su versatilidad les ha permitido convertirse en los miembros más ricos en especies de todos los gremios ecológicos en la mayoría de los entornos.
Los artrópodos marinos existentes varían en tamaño desde el Stygotantulus un crustáceo microscópico hasta el cangrejo gigante japonés. La cavidad interna primaria de los artrópodos es un hemocoel, que aloja sus órganos internos, y a través del cual circula su hemolinfa, análogo de la sangre; tienen sistemas circulatorios abiertos. Al igual que sus órganos exteriores, los órganos internos de los artrópodos generalmente están construidos con segmentos repetidos. Su sistema nervioso es "similar a una escalera", con cuerdas nerviosas ventrales pareadas que recorren todos los segmentos y forman ganglios pareados en cada segmento. Sus cabezas están formadas por la fusión de un número variable de segmentos, y sus cerebros están formados por la fusión de los ganglios de estos segmentos y rodean el esófago. Los sistemas respiratorios y excretores de los artrópodos varían, dependiendo tanto de su entorno como del subfilo al que pertenecen.
Su visión se basa en varias combinaciones de ojos compuestos y ocelos de pigmento: en la mayoría de las especies, los ocelos solo pueden detectar la dirección desde la que proviene la luz, y los ojos compuestos son la principal fuente de información. Los artrópodos también tienen una amplia gama de sensores químicos y mecánicos, en su mayoría basados en modificaciones de las muchas setas (cerdas) que se proyectan a través de sus cutículas. Los métodos de reproducción y desarrollo de los artrópodos son diversos; todas las especies terrestres utilizan la fertilización interna, pero esto a menudo se realiza mediante la transferencia indirecta de los espermatozoides a través de un apéndice o el suelo, en lugar de por inyección directa. Todas las especies marinas ponen huevos y usan fertilización interna o externa. Las crías de artrópodos varían de adultos en miniatura a larvas que carecen de miembros articulados y eventualmente experimentan una metamorfosis total para producir la forma adulta.
-
![Los trilobites, ya extintos, poblaron los océanos durante 270 millones de años.[107]](//upload.wikimedia.org/wikipedia/commons/thumb/0/08/Olenoides_typicalis.jpg/114px-Olenoides_typicalis.jpg) Los trilobites, ya extintos, poblaron los océanos durante 270 millones de años.[107]
Los trilobites, ya extintos, poblaron los océanos durante 270 millones de años.[107] -
 El cangrejo herradura es un fósil viviente, que ha permanecido inalterado durante 450 millones de años
El cangrejo herradura es un fósil viviente, que ha permanecido inalterado durante 450 millones de años -
![El mayor de los artrópodos conocidos es el escorpión marino Jaekelopterus rhenaniae, que vivió hace 390 millones de años. Media 2.5 m de largo.[108][109]](//upload.wikimedia.org/wikipedia/commons/thumb/a/af/Jaekelopterus_rhenaniae_reconstruction.jpg/265px-Jaekelopterus_rhenaniae_reconstruction.jpg) El mayor de los artrópodos conocidos es el escorpión marino Jaekelopterus rhenaniae, que vivió hace 390 millones de años. Media 2.5 m de largo.[108][109]
El mayor de los artrópodos conocidos es el escorpión marino Jaekelopterus rhenaniae, que vivió hace 390 millones de años. Media 2.5 m de largo.[108][109]
![Los trilobites, ya extintos, poblaron los océanos durante 270 millones de años.[107]](/wiki/Archivo:Olenoides_typicalis.jpg)
_rotated.jpg)
![El mayor de los artrópodos conocidos es el escorpión marino Jaekelopterus rhenaniae, que vivió hace 390 millones de años. Media 2.5 m de largo.[108][109]](/wiki/Archivo:Jaekelopterus_rhenaniae_reconstruction.jpg)
- Crustáceos
-
 Muchos crustáceos son muy pequeños, como este diminuto amfipodo, son una parte importante del zooplancton de los océanos
Muchos crustáceos son muy pequeños, como este diminuto amfipodo, son una parte importante del zooplancton de los océanos -
![La langosta mantis posee los ojos más desarrollados del mundo animal,[110] y abaten a sus presas revoleando sus tenazas que se asemejan a mazas.[111]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b2/OdontodactylusScyllarus.jpg/239px-OdontodactylusScyllarus.jpg) La langosta mantis posee los ojos más desarrollados del mundo animal,[110] y abaten a sus presas revoleando sus tenazas que se asemejan a mazas.[111]
La langosta mantis posee los ojos más desarrollados del mundo animal,[110] y abaten a sus presas revoleando sus tenazas que se asemejan a mazas.[111] -
![El cangrejo gigante de Tasmania tiene una larga vida y crece lentamente, lo cual lo hace vulnerable a la pesca.[112]](//upload.wikimedia.org/wikipedia/commons/thumb/2/2c/J_J_Wild_Pseudocarcinus_cropped.jpg/264px-J_J_Wild_Pseudocarcinus_cropped.jpg) El cangrejo gigante de Tasmania tiene una larga vida y crece lentamente, lo cual lo hace vulnerable a la pesca.[112]
El cangrejo gigante de Tasmania tiene una larga vida y crece lentamente, lo cual lo hace vulnerable a la pesca.[112] -
![Cigala.[113]](//upload.wikimedia.org/wikipedia/commons/thumb/b/b1/Nephrops_norvegicus.jpg/240px-Nephrops_norvegicus.jpg) Cigala.[113]
Cigala.[113]

![La langosta mantis posee los ojos más desarrollados del mundo animal,[110] y abaten a sus presas revoleando sus tenazas que se asemejan a mazas.[111]](/wiki/Archivo:OdontodactylusScyllarus.jpg)
![El cangrejo gigante de Tasmania tiene una larga vida y crece lentamente, lo cual lo hace vulnerable a la pesca.[112]](/wiki/Archivo:J_J_Wild_Pseudocarcinus_cropped.jpg)
![Cigala.[113]](/wiki/Archivo:Nephrops_norvegicus.jpg)
Vertebrados editar
Peces editar
La anatomía de los peces comprende un corazón con dos cámaras, opérculo, vejiga natatoria, escamas, ojos adaptados para ver bajo el agua, y células secretorias que producen moco. Los peces respiran extrayendo oxígeno del agua mediante branquias. Las aletas le permiten propulsarse y controlar su estabilidad en el agua. Los peces se organizan en dos grandes grupos: peces con esqueletos óseos y peces con esqueletos cartilaginosos. Existen más de 33 000 especies de peces,[114] de las cuales unas 20 000 corresponden a peces marinos.[115]
Peces con esqueleto óseo editar
Los peces con esqueletos óseos poseen mandíbula y esqueletos de hueso en vez de cartílago. Estos peces poseen una placa ósea que cubre sus branquias que se denomina opérculo, y pueden tener distintas espinas en sus aletas. Aproximadamente el 90% de los peces poseen esqueleto óseo.
El esqueleto está compuesto por el esqueleto axial (columna vertebral), el esqueleto cefálico (cabeza), el esqueleto zonal (las dos cinturas que se anclan a las aletas pelvianas y torácicas respectivamente) y el esqueleto apendicular (aletas).
Presentan mayoritariamente boca terminal, capaz de realizar movimientos muy precisos gracias a que poseen huesos dérmicos articulados. Los dientes salen de algunos de estos huesos dérmicos y carecen de reemplazo cuando se caen o rompen. Además, su aleta caudal es homocerca. Aparte de su esqueleto interno calcificado u osificado (la excepción es el esturión, que lo tiene cartilaginoso como los condrictios), también poseen huesos en la dermis de su tegumento en forma de escamas, lo que se conoce como dermatoesqueleto. Estas escamas tienen un importante valor taxonómico, ya que el tipo y número (especialmente en la línea lateral y transversalmente) son usados como rasgos identificativos de los distintos grupos.
-
 Thunnus thynnus (Atún rojo)
Thunnus thynnus (Atún rojo) -
 Carassius carassius (Carpín)
Carassius carassius (Carpín) -
 Coelacanthimorpha (Celacanto)
Coelacanthimorpha (Celacanto) -
 El extinto Leedsichthys es el mayor de los peces con esqueleto óseo conocidos
El extinto Leedsichthys es el mayor de los peces con esqueleto óseo conocidos -
 Salmo salar (Salmón del Atlántico)
Salmo salar (Salmón del Atlántico) -
 Xiphias gladius (Pez espada)
Xiphias gladius (Pez espada)






Peces con esqueleto cartilaginoso editar
Los peces cartilaginosos, tales como los tiburones y rayas, poseen mandíbulas y esqueletos conformados a base de cartílago en vez de hueso. El megalodon es una especie extinta de tiburón que vivió hace unos 1,5 millones de años. Se parecía mucho a una versión más robusta del gran tiburón blanco, pero era mucho más largo con longitudes que alcanzaban 20 m.[116] Habitaba en todos los océanos[117] y fue el más grande y poderosos de los depredadores en la historia de los vertebrados,[116] y probablemente tenía un impacto importante en la vida marina.[118] El tiburón de Groenlandia es el vertebrado que vive más años llegando a vivir 400 años.[119]
-
 Es posible que los peces cartilaginosos hayan evolucionado a partir de los tiburones espinosos
Es posible que los peces cartilaginosos hayan evolucionado a partir de los tiburones espinosos -
 El megalodon, ya extinto, se asemeja a un gran tiburón blanco
El megalodon, ya extinto, se asemeja a un gran tiburón blanco -
 El tiburón de Groenlandia es el vertebrado que posee una vida más larga
El tiburón de Groenlandia es el vertebrado que posee una vida más larga -

-
![Los peces sierra son rayas con extensas rostrums que se asemejan a una sierra.[120]](//upload.wikimedia.org/wikipedia/commons/thumb/9/9b/Sawfish_genova.jpg/200px-Sawfish_genova.jpg)
-
![La manta raya gigante, que es la mayor raya del mundo, fue objeto de persecución por la industria pesquera y en la actualidad es una especie vulnerable.[121]](//upload.wikimedia.org/wikipedia/commons/thumb/8/85/Manta_alfredi_fushivaru_thila.jpg/200px-Manta_alfredi_fushivaru_thila.jpg) La manta raya gigante, que es la mayor raya del mundo, fue objeto de persecución por la industria pesquera y en la actualidad es una especie vulnerable.[121]
La manta raya gigante, que es la mayor raya del mundo, fue objeto de persecución por la industria pesquera y en la actualidad es una especie vulnerable.[121]




![Los peces sierra son rayas con extensas rostrums que se asemejan a una sierra.[120]](/wiki/Archivo:Sawfish_genova.jpg)
![La manta raya gigante, que es la mayor raya del mundo, fue objeto de persecución por la industria pesquera y en la actualidad es una especie vulnerable.[121]](/wiki/Archivo:Manta_alfredi_fushivaru_thila.jpg)
Peces sin mandíbula editar
Los mixinos forman una clase de aproximadamente 20 especies de peces marinos productores de moco en forma de anguila. Son los únicos animales vivos conocidos que tienen un cráneo pero no una columna vertebral. Las lampreas forman una superclase que contiene 38 especies conocidas de peces sin mandíbulas. La lamprea adulta se caracteriza por una boca de succión con forma de embudo dentada. Aunque son bien conocidos por escarbar en la carne de otros peces para chupar su sangre, solo 18 especies de lampreas son en realidad parásitas. Juntos, los mixinos y las lampreas son el grupo hermano de los vertebrados. El mixino existente en la actualidad es muy similar al mixino de hace 300 millones de años. Las lampreas son un linaje muy antiguo de vertebrados, aunque su relación exacta con los mixinos y los vertebrados con mandíbulas aún es un tema en disputa.
-

-
 A menudo las lampreas son parasitarias y poseen una boca con dientes y que asemeja un embudo para succionar
A menudo las lampreas son parasitarias y poseen una boca con dientes y que asemeja un embudo para succionar -
 El extinto Pteraspidomorphi, un ancestro de los vertebrados con mandíbula
El extinto Pteraspidomorphi, un ancestro de los vertebrados con mandíbula



Plancton editar

El plancton comprende un grupo diverso de organismos que viven en los cuerpos de agua y que no pueden nadar contra la corriente.[122] Ellos son una fuente de alimento crucial de numerosos organismos acuáticos más grandes, tales como peces y ballenas.
Estos organismos abarcan bacterias, archaea, algas, protozoos y animales que flotan o son arrastrados por las corrientes donde habitan, por ejemplo, la zona pelágica de océanos, mares, o cuerpos de agua dulce. Esencialmente, los tipos de plancton se definen por sus nichos ecológicos en vez de mediante clasificación filogenética o taxonómica.
Si bien numerosas especies planctónicas poseen un tamaño microscópico, el plancton comprende organismos que abarcan una amplia gama de tamaños, incluidos organismos grandes tales como medusas.[123]
El rol e importancia del fitoplancton pueden ser comprendidos si se observa la posición crítica que ocupan como los productores primarios más numerosos de la Tierra. El fitoplancton se categoriza en cyanobacteria (también denominadas algas/bacterias azul-verde), varios tipos de algas (rojas, verdes, marrones y amarillo-verdosas), diatomeas, dinoflagelados, euglenoidos, cocolitóforos, criptomonados, crisofitos, clorófitos, prasinofitas, y silicoflagelados.
- Fitoplancton
-
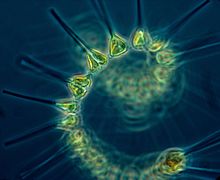 El fitoplancton es la base de la cadena alimentaria en los océanos.
El fitoplancton es la base de la cadena alimentaria en los océanos. -
 Existen de muchas formas y tamaños.
Existen de muchas formas y tamaños. -
 Las diatomeas son unos de los tipos más comunes de fitoplancton
Las diatomeas son unos de los tipos más comunes de fitoplancton -
 Espuma verde de cyanobacteria arrojada por el oleaje sobre una roca en California
Espuma verde de cyanobacteria arrojada por el oleaje sobre una roca en California -
 Proliferación de algas frente a la costa en el sur de Inglaterra.
Proliferación de algas frente a la costa en el sur de Inglaterra.




El zooplancton tiende a ser más grande, y no todos son microscópicos. Muchos protozoos son zooplancton, incluidos los dinoflagelados, zooflagelados, foraminiferanos, y radiolarios. Algunos de ellos (tales como los dinoflagelados) también son fitoplancton; la distinción entre plantas y animales a menudo deja de ser aplicable en organismos muy pequeños. Otro zooplancton comprende cnidarios, ctenofóros, quetognatos, moluscos, artrópodos, urocordados, y anélidos tales como poliquetos.
- Zooplancton
-
 Anfipodo marino
Anfipodo marino -

-

-
![Tomopteris, un gusano planctonico segmentado con bioluminescencia[124]](//upload.wikimedia.org/wikipedia/commons/thumb/4/40/Tomopteriskils.jpg/265px-Tomopteriskils.jpg)


_edit.jpg)
![Tomopteris, un gusano planctonico segmentado con bioluminescencia[124]](/wiki/Archivo:Tomopteriskils.jpg)
Muchos animales más grandes comienzan su existencia como zooplancton antes de crecer y adoptar sus formas adultas.
- Freza, larvas y juveniles
-

-
 Macho de coral estrella liberando esperma al agua
Macho de coral estrella liberando esperma al agua -
 Larva de pez luna
Larva de pez luna -

-
 Juvenil planctónico de calamar
Juvenil planctónico de calamar





Notas editar
- ↑ Esta es la medida tomada por el buque Kaikō en marzo de 1995 y se considera la medición más precisa hasta la fecha. Vea el artículo de el Abismo de Challenger para más detalles.
Referencias editar
- ↑ Living Ocean «Living Ocean». NASA Science (en inglés). Consultado el 17 de diciembre de 2016.
- ↑ Drogin, Bob (2 de agosto de 2009). «Mapping an ocean of species». Los Angeles Times. Consultado el 18 de agosto de 2009.
- ↑ Paul, Gregory S. (2010). The Evolution of Dinosaurs and their World. «The Princeton Field Guide to Dinosaurs». Princeton University Press (Princeton). p. 19.
- ↑ Bortolotti, Dan (2008). «Wild Blue: A Natural History of the World's Largest Animal». St. Martin's Press (en inglés).
- ↑ Agre, P (2006). «The aquaporin water channels». Proceedings of the American Thoracic Society 3 (1). pp. 5-13. PMC 2658677. PMID 16493146. doi:10.1513/pats.200510-109JH.
- ↑ Collins J. C. (1991) The Matrix of Life: A View of Natural Molecules from the Perspective of Environmental Water Molecular Presentations. ISBN 9780962971907.
- ↑ «7,000 m Class Remotely Operated Vehicle KAIKO 7000» (en inglés). Japan Agency for Marine-Earth Science and Technology (JAMSTEC). Archivado desde el original el 10 de abril de 2020. Consultado el 7 de junio de 2008.
- ↑ Charette, Matthew A.; Smith, Walter H. F. (Junio de 2010). «The Volume of Earth's Ocean». Oceanography (en inglés) 23 (2): 112-14. doi:10.5670/oceanog.2010.51. Archivado desde el original el 2 de agosto de 2013. Consultado el 6 de junio de 2013.
- ↑ «sphere depth of the ocean – hydrology». Encyclopædia Britannica. Consultado el 12 de abril de 2015.
- ↑ «Third rock from the Sun – restless Earth». NASA's Cosmos (en inglés). Consultado el 12 de abril de 2015.
- ↑ Perlman, Howard (17 de marzo de 2014). «The World's Water». USGS Water-Science School. Consultado el 12 de abril de 2015.
- ↑ Kennish, Michael J. (2001). Practical handbook of marine science. Marine science series (en inglés) (3ra edición). CRC Press. p. 35. ISBN 0-8493-2391-6.
- ↑ «Age of the Earth». United States Geological Survey. 9 de julio de 2007. Consultado el 31 de mayo de 2015.
- ↑ Dalrymple, 2001, pp. 205-221
- ↑ Manhesa, Gérard; Allègre, Claude J.; Dupréa, Bernard; Hamelin, Bruno (May 1980). «Lead isotope study of basic-ultrabasic layered complexes: Speculations about the age of the earth and primitive mantle characteristics». Earth and Planetary Science Letters (Amsterdam, the Netherlands: Elsevier) 47 (3): 370-382. Bibcode:1980E&PSL..47..370M. ISSN 0012-821X. doi:10.1016/0012-821X(80)90024-2.
- ↑ Schopf, J. William; Kudryavtsev, Anatoliy B.; Czaja, Andrew D.; Tripathi, Abhishek B. (5 de octubre de 2007). «Evidence of Archean life: Stromatolites and microfossils». Precambrian Research (en inglés) (Amsterdam, the Netherlands: Elsevier) 158 (3-4): 141-155. ISSN 0301-9268. doi:10.1016/j.precamres.2007.04.009.
- ↑ Raven y Johnson, 2002, p. 68
- ↑ Borenstein, Seth (13 de noviembre de 2013). «Oldest fossil found: Meet your microbial mom». Excite (Yonkers, N.Y.: Mindspark Interactive Network). Associated Press. Consultado el 31 de mayo de 2015.
- ↑ Pearlman, Jonathan (13 de noviembre de 2013). «'Oldest signs of life on Earth found'». The Daily Telegraph (en inglés) (Londres: Telegraph Media Group). Consultado el 15 de diciembre de 2014.
- ↑ Noffke, Nora; Christian, Daniel; Wacey, David; Hazen, Robert M. (16 de noviembre de 2013). «Microbially Induced Sedimentary Structures Recording an Ancient Ecosystem in the ca. 3.48 Billion-Year-Old Dresser Formation, Pilbara, Western Australia». Astrobiology (en inglés) (New Rochelle, N.Y.: Mary Ann Liebert, Inc.) 13 (12): 1103-1124. Bibcode:2013AsBio..13.1103N. ISSN 1531-1074. PMC 3870916. PMID 24205812. doi:10.1089/ast.2013.1030.
- ↑ Ohtomo, Yoko; Kakegawa, Takeshi; Ishida, Akizumi et al. (January 2014). «Evidence for biogenic graphite in early Archaean Isua metasedimentary rocks». Nature Geoscience (London: Nature Publishing Group) 7 (1): 25-28. Bibcode:2014NatGe...7...25O. ISSN 1752-0894. doi:10.1038/ngeo2025.
- ↑ a b Borenstein, Seth (19 de octubre de 2015). «Hints of life on what was thought to be desolate early Earth». Excite (en inglés) (Yonkers, N.Y.: Mindspark Interactive Network). Associated Press. Consultado el 20 de octubre de 2015.
- ↑ Bell, Elizabeth A.; Boehnike, Patrick; Harrison, T. Mark et al. (24 de noviembre de 2015). «Potentially biogenic carbon preserved in a 4.1 billion-year-old zircon» (PDF). Proceedings of the National Academy of Sciences of the United States of America (en inglés) (Washington, D.C.: National Academy of Sciences) 112 (47): 14518-14521. ISSN 0027-8424. PMC 4664351. PMID 26483481. doi:10.1073/pnas.1517557112. Consultado el 30 de diciembre de 2015.
- ↑ Penny, David; Poole, Anthony (December 1999). «The nature of the last universal common ancestor». Current Opinion in Genetics & Development (Amsterdam, the Netherlands: Elsevier) 9 (6): 672-677. ISSN 0959-437X. PMID 10607605. doi:10.1016/S0959-437X(99)00020-9.
- ↑ Theobald, Douglas L. (13 de mayo de 2010). «A formal test of the theory of universal common ancestry». Nature (London: Nature Publishing Group) 465 (7295): 219-222. Bibcode:2010Natur.465..219T. ISSN 0028-0836. PMID 20463738. doi:10.1038/nature09014.
- ↑ Doolittle, W. Ford (February 2000). «Uprooting the Tree of Life» (PDF). Scientific American (Stuttgart: Georg von Holtzbrinck Publishing Group) 282 (2): 90-95. ISSN 0036-8733. PMID 10710791. doi:10.1038/scientificamerican0200-90. Archivado desde el original el 7 de septiembre de 2006. Consultado el 5 de abril de 2015.
- ↑ Peretó, Juli (March 2005). «Controversies on the origin of life» (PDF). International Microbiology (Barcelona: Spanish Society for Microbiology) 8 (1): 23-31. ISSN 1139-6709. PMID 15906258. Archivado desde el original el 24 de agosto de 2015.
- ↑ Joyce, Gerald F. (11 de julio de 2002). «The antiquity of RNA-based evolution». Nature (London: Nature Publishing Group) 418 (6894): 214-221. Bibcode:2002Natur.418..214J. ISSN 0028-0836. PMID 12110897. doi:10.1038/418214a.
- ↑ Trevors, Jack T.; Psenner, Roland (December 2001). «From self-assembly of life to present-day bacteria: a possible role for nanocells». FEMS Microbiology Reviews (Amsterdam, the Netherlands: Elsevier on behalf of the Federation of European Microbiological Societies) 25 (5): 573-582. ISSN 1574-6976. PMID 11742692. doi:10.1111/j.1574-6976.2001.tb00592.x.
- ↑ Bapteste, Eric; Walsh, David A. (June 2005). «Does the 'Ring of Life' ring true?». Trends in Microbiology (Cambridge, Mass.: Cell Press) 13 (6): 256-261. ISSN 0966-842X. PMID 15936656. doi:10.1016/j.tim.2005.03.012.
- ↑ Darwin, 1859, p. 1
- ↑ Doolittle, W. Ford; Bapteste, Eric (13 de febrero de 2007). «Pattern pluralism and the Tree of Life hypothesis». Proceedings of the National Academy of Sciences of the United States of America (Washington, D.C.: National Academy of Sciences) 104 (7): 2043-2049. Bibcode:2007PNAS..104.2043D. ISSN 0027-8424. PMC 1892968. PMID 17261804. doi:10.1073/pnas.0610699104.
- ↑ Kunin, Victor; Goldovsky, Leon; Darzentas, Nikos; Ouzounis, Christos A. (July 2005). «The net of life: Reconstructing the microbial phylogenetic network». Genome Research (Cold Spring Harbor, N.Y.: Cold Spring Harbor Laboratory Press) 15 (7): 954-959. ISSN 1088-9051. PMC 1172039. PMID 15965028. doi:10.1101/gr.3666505.
- ↑ Jablonski, David (25 de junio de 1999). «The Future of the Fossil Record». Science (Washington, D.C.: American Association for the Advancement of Science) 284 (5423): 2114-2116. ISSN 0036-8075. PMID 10381868. doi:10.1126/science.284.5423.2114.
- ↑ Mason, Stephen F. (6 de septiembre de 1984). «Origins of biomolecular handedness». Nature (London: Nature Publishing Group) 311 (5981): 19-23. Bibcode:1984Natur.311...19M. ISSN 0028-0836. PMID 6472461. doi:10.1038/311019a0.
- ↑ Wolf, Yuri I.; Rogozin, Igor B.; Grishin, Nick V.; Koonin, Eugene V. (1 de septiembre de 2002). «Genome trees and the tree of life». Trends in Genetics (Cambridge, Mass.: Cell Press) 18 (9): 472-479. ISSN 0168-9525. PMID 12175808. doi:10.1016/S0168-9525(02)02744-0.
- ↑ Varki, Ajit; Altheide, Tasha K. (December 2005). «Comparing the human and chimpanzee genomes: searching for needles in a haystack». Genome Research (Cold Spring Harbor, N.Y.: Cold Spring Harbor Laboratory Press) 15 (12): 1746-1758. ISSN 1088-9051. PMID 16339373. doi:10.1101/gr.3737405.
- ↑ a b Cavalier-Smith, Thomas (29 de junio de 2006). «Cell evolution and Earth history: stasis and revolution». Philosophical Transactions of the Royal Society B: Biological Sciences (London: Royal Society) 361 (1470): 969-1006. ISSN 0962-8436. PMC 1578732. PMID 16754610. doi:10.1098/rstb.2006.1842.
- ↑ Schopf, J. William (29 de junio de 2006). «Fossil evidence of Archaean life». Philosophical Transactions of the Royal Society B: Biological Sciences (London: Royal Society) 361 (1470): 869-885. ISSN 0962-8436. PMC 1578735. PMID 16754604. doi:10.1098/rstb.2006.1834.
- Altermann, Wladyslaw; Kazmierczak, Józef (November 2003). «Archean microfossils: a reappraisal of early life on Earth». Research in Microbiology (Amsterdam, the Netherlands: Elsevier for the Pasteur Institute) 154 (9): 611-617. ISSN 0923-2508. PMID 14596897. doi:10.1016/j.resmic.2003.08.006.
- ↑ Schopf, J. William (19 de julio de 1994). «Disparate rates, differing fates: tempo and mode of evolution changed from the Precambrian to the Phanerozoic». Proceedings of the National Academy of Sciences of the United States of America (Washington, D.C.: National Academy of Sciences) 91 (15): 6735-6742. Bibcode:1994PNAS...91.6735S. ISSN 0027-8424. PMC 44277. PMID 8041691. doi:10.1073/pnas.91.15.6735.
- ↑ Poole, Anthony M.; Penny, David (January 2007). «Evaluating hypotheses for the origin of eukaryotes». BioEssays (Hoboken, N.J.: John Wiley & Sons) 29 (1): 74-84. ISSN 0265-9247. PMID 17187354. doi:10.1002/bies.20516.
- ↑ Dyall, Sabrina D.; Brown, Mark T.; Johnson, Patricia J. (9 de abril de 2004). «Ancient Invasions: From Endosymbionts to Organelles». Science (Washington, D.C.: American Association for the Advancement of Science) 304 (5668): 253-257. Bibcode:2004Sci...304..253D. ISSN 0036-8075. PMID 15073369. doi:10.1126/science.1094884.
- ↑ Martin, William (October 2005). «The missing link between hydrogenosomes and mitochondria». Trends in Microbiology (Cambridge, Mass.: Cell Press) 13 (10): 457-459. ISSN 0966-842X. PMID 16109488. doi:10.1016/j.tim.2005.08.005.
- ↑ Lang, B. Franz; Gray, Michael W.; Burger, Gertraud (December 1999). «Mitochondrial genome evolution and the origin of eukaryotes». Annual Review of Genetics (Palo Alto, Calif.: Annual Reviews) 33: 351-397. ISSN 0066-4197. PMID 10690412. doi:10.1146/annurev.genet.33.1.351.
- ↑ DeLong, Edward F.; Pace, Norman R. (1 de agosto de 2001). «Environmental Diversity of Bacteria and Archaea». Systematic Biology (Oxford: Oxford University Press on behalf of the Society of Systematic Biologists) 50 (4): 470-478. ISSN 1063-5157. PMID 12116647. doi:10.1080/106351501750435040.
- ↑ Kaiser, Dale (December 2001). «Building a multicellular organism». Annual Review of Genetics (Palo Alto, Calif.: Annual Reviews) 35: 103-123. ISSN 0066-4197. PMID 11700279. doi:10.1146/annurev.genet.35.102401.090145.
- ↑ Zimmer, Carl (7 de enero de 2016). «Genetic Flip Helped Organisms Go From One Cell to Many». The New York Times. Consultado el 7 de enero de 2016.
- ↑ Valentine, James W.; Jablonski, David; Erwin, Douglas H. (1 de marzo de 1999). «Fossils, molecules and embryos: new perspectives on the Cambrian explosion». Development (Cambridge: The Company of Biologists) 126 (5): 851-859. ISSN 0950-1991. PMID 9927587. Consultado el 30 de diciembre de 2014.
- ↑ Ohno, Susumu (January 1997). «The reason for as well as the consequence of the Cambrian explosion in animal evolution». Journal of Molecular Evolution (New York: Springer-Verlag New York) 44 (Suppl. 1): S23-S27. ISSN 0022-2844. PMID 9071008. doi:10.1007/PL00000055.
- Valentine, James W.; Jablonski, David (2003). «Morphological and developmental macroevolution: a paleontological perspective». The International Journal of Developmental Biology (Bilbao, Spain: University of the Basque Country Press) 47 (7-8): 517-522. ISSN 0214-6282. PMID 14756327. Consultado el 30 de diciembre de 2014.
- ↑ Wellman, Charles H.; Osterloff, Peter L.; Mohiuddin, Uzma (2003). «Fragments of the earliest land plants». Nature 425 (6955): 282-285. Bibcode:2003Natur.425..282W. PMID 13679913. doi:10.1038/nature01884.
- ↑ Barton, Nicholas (2007). Evolution. pp. 273-274. ISBN 9780199226320. Consultado el 30 de septiembre de 2012.
- ↑ Waters, Elizabeth R. (December 2003). «Molecular adaptation and the origin of land plants». Molecular Phylogenetics and Evolution (Academic Press) 29 (3): 456-463. ISSN 1055-7903. PMID 14615186. doi:10.1016/j.ympev.2003.07.018.
- ↑ Mayhew, Peter J. (August 2007). «Why are there so many insect species? Perspectives from fossils and phylogenies». Biological Reviews (Cambridge: Cambridge University Press on behalf of the Cambridge Philosophical Society) 82 (3): 425-454. ISSN 1464-7931. PMID 17624962. doi:10.1111/j.1469-185X.2007.00018.x.
- ↑ Carroll, Robert L. (May 2007). «The Palaeozoic Ancestry of Salamanders, Frogs and Caecilians». Zoological Journal of the Linnean Society (Hoboken, N.J.: Wiley-Blackwell) 150 (Supplement s1): 1-140. ISSN 1096-3642. doi:10.1111/j.1096-3642.2007.00246.x.
- ↑ Wible, John R.; Rougier, Guillermo W.; Novacek, Michael J.; Asher, Robert J. (21 de junio de 2007). «Cretaceous eutherians and Laurasian origin for placental mammals near the K/T boundary». Nature (London: Nature Publishing Group) 447 (7147): 1003-1006. Bibcode:2007Natur.447.1003W. ISSN 0028-0836. PMID 17581585. doi:10.1038/nature05854.
- ↑ Witmer, Lawrence M. (28 de julio de 2011). «Palaeontology: An icon knocked from its perch». Nature (London: Nature Publishing Group) 475 (7357): 458-459. ISSN 0028-0836. PMID 21796198. doi:10.1038/475458a.
- ↑ Schloss, Patrick D.; Handelsman, Jo (December 2004). «Status of the Microbial Census». Microbiology and Molecular Biology Reviews (Washington, D.C.: American Society for Microbiology) 68 (4): 686-691. ISSN 1092-2172. PMC 539005. PMID 15590780. doi:10.1128/MMBR.68.4.686-691.2004.
- ↑ Miller y Spoolman, 2012, p. 62
- ↑ Mora, Camilo; Tittensor, Derek P.; Adl, Sina et al. (23 de agosto de 2011). «How Many Species Are There on Earth and in the Ocean?». PLoS Biology (San Francisco: Public Library of Science) 9 (8): e1001127. ISSN 1545-7885. PMC 3160336. PMID 21886479. doi:10.1371/journal.pbio.1001127.
- ↑ Yinon M. Bar-On; Rob Phillips; Ron Milo (Junio de 2018). «The biomass distribution on Earth». Proceedings of the National Academy of Sciences of the United States of America (en inglés) 115 (25): 6506-6511. Bibcode:2018PNAS..115.6506B. PMC 6016768. PMID 29784790. doi:10.1073/pnas.1711842115.
- ↑ Madigan M; Martinko J, eds. (2006). Brock Biology of Microorganisms (13ª edición). Pearson Education. ISBN 978-0-321-73551-5.
- ↑ Ed Rybicki (1990). «The classification of organisms at the edge of life, or problems with virus systematics». South African Journal of Science 86: 182-6. ISSN 0038-2353.
- ↑ Lwoff A (1956). «The concept of virus». Journal of General Microbiology 17 (2): 239-53. PMID 13481308. doi:10.1099/00221287-17-2-239.
- ↑ 2002 WHO mortality data Accessed 20 January 2007
- ↑ «Functions of global ocean microbiome key to understanding environmental changes». www.sciencedaily.com. University of Georgia. 10 de diciembre de 2015. Consultado el 11 de diciembre de 2015.
- ↑ Suttle, C.A. (2005). «Viruses in the Sea». Nature 437 (9): 356-361. Bibcode:2005Natur.437..356S. PMID 16163346. S2CID 4370363. doi:10.1038/nature04160.
- ↑ Shors p. 5
- ↑ Shors p. 593
- ↑ Curtis A. Suttle (Octubre de 2007). «Marine viruses — major players in the global ecosystem». Nature Reviews. Microbiology 5 (10): 801-12. PMID 17853907. S2CID 4658457. doi:10.1038/nrmicro1750.
- ↑ Living Bacteria Are Riding Earth’s Air Currents Smithsonian Magazine, 11 January 2016.
- ↑ Robbins, Jim (13 de abril de 2018). «Trillions Upon Trillions of Viruses Fall From the Sky Each Day». The New York Times. Consultado el 14 de abril de 2018.
- ↑ Reche, Isabel; D’Orta, Gaetano; Mladenov, Natalie; Winget, Danielle M; Suttle, Curtis A (29 de enero de 2018). «Deposition rates of viruses and bacteria above the atmospheric boundary layer». ISME Journal 12 (4): 1154-1162. PMC 5864199. PMID 29379178. doi:10.1038/s41396-017-0042-4.
- ↑ Staff (2014). «The Biosphere». Aspen Global Change Institute. Archivado desde el original el 2 de septiembre de 2010. Consultado el 10 de noviembre de 2014.
- ↑ a b c Choi, Charles Q. (17 de marzo de 2013). «Microbes Thrive in Deepest Spot on Earth». LiveScience. Consultado el 17 de marzo de 2013.
- ↑ Glud, Ronnie; Wenzhöfer, Frank; Middelboe, Mathias; Oguri, Kazumasa; Turnewitsch, Robert; Canfield, Donald E.; Kitazato, Hiroshi (17 de marzo de 2013). «High rates of microbial carbon turnover in sediments in the deepest oceanic trench on Earth». Nature Geoscience 6 (4): 284-288. Bibcode:2013NatGe...6..284G. doi:10.1038/ngeo1773.
- ↑ Oskin, Becky (14 de marzo de 2013). «Intraterrestrials: Life Thrives in Ocean Floor». LiveScience. Consultado el 17 de marzo de 2013.
- ↑ Morelle, Rebecca (15 de diciembre de 2014). «Microbes discovered by deepest marine drill analysed». BBC News. Consultado el 15 de diciembre de 2014.
- ↑ Takai K; Nakamura K; Toki T; Tsunogai U et al. (2008). «Cell proliferation at 122°C and isotopically heavy CH4 production by a hyperthermophilic methanogen under high-pressure cultivation». Proceedings of the National Academy of Sciences of the United States of America 105 (31): 10949-54. Bibcode:2008PNAS..10510949T. PMC 2490668. PMID 18664583. doi:10.1073/pnas.0712334105.
- ↑ Fox, Douglas (20 de agosto de 2014). «Lakes under the ice: Antarctica's secret garden». Nature 512 (7514): 244-246. Bibcode:2014Natur.512..244F. PMID 25143097. doi:10.1038/512244a.
- ↑ Mack, Eric (20 de agosto de 2014). «Life Confirmed Under Antarctic Ice; Is Space Next?». Forbes. Consultado el 21 de agosto de 2014.
- ↑ Whittaker, R.H.; Margulis, L. (1978). «Protist classification and the kingdoms of organisms». Biosystems 10 (1–2): 3-18. PMID 418827. doi:10.1016/0303-2647(78)90023-0.
- ↑ Faure, E; Not, F; Benoiston, AS; Labadie, K; Bittner, L; Ayata, SD (2019). «Mixotrophic protists display contrasted biogeographies in the global ocean». ISME Journal 13 (4): 1072-1083. PMC 6461780. PMID 30643201. doi:10.1038/s41396-018-0340-5.
- ↑ a b Leles, S.G.; Mitra, A.; Flynn, K.J.; Stoecker, D.K.; Hansen, P.J.; Calbet, A.; McManus, G.B.; Sanders, R.W.; Caron, D.A.; Not, F.; Hallegraeff, G.M. (1860). «Oceanic protists with different forms of acquired phototrophy display contrasting biogeographies and abundance». Proceedings of the Royal Society B: Biological Sciences 284 (1860): 20170664. PMC 5563798. PMID 28768886. doi:10.1098/rspb.2017.0664.
- ↑ Budd, Graham E; Jensen, Sören (2017). «The origin of the animals and a 'Savannah' hypothesis for early bilaterian evolution». Biological Reviews 92 (1): 446-473. PMID 26588818. doi:10.1111/brv.12239.
- ↑ Cavalier-Smith T (December 1993). «Kingdom protozoa and its 18 phyla». Microbiological Reviews 57 (4): 953-94. PMC 372943. PMID 8302218. doi:10.1128/MMBR.57.4.953-994.1993.
- ↑ Corliss JO (1992). «Should there be a separate code of nomenclature for the protists?». BioSystems 28 (1–3): 1-14. PMID 1292654. doi:10.1016/0303-2647(92)90003-H.
- ↑ Slapeta J, Moreira D, López-García P (2005). «The extent of protist diversity: insights from molecular ecology of freshwater eukaryotes». Proceedings of the Royal Society B: Biological Sciences 272 (1576): 2073-81. PMC 1559898. PMID 16191619. doi:10.1098/rspb.2005.3195.
- ↑ Moreira D, López-García P (2002). «The molecular ecology of microbial eukaryotes unveils a hidden world». Trends in Microbiology 10 (1): 31-8. PMID 11755083. doi:10.1016/S0966-842X(01)02257-0.
- ↑ The Air You're Breathing? A Diatom Made That
- ↑ «More on Diatoms». University of California Museum of Paleontology. Archivado desde el original el 4 de octubre de 2012. Consultado el 27 de junio de 2019.
- ↑ Janet Fang (6 de abril de 2010). «Animals thrive without oxygen at sea bottom». Nature 464 (7290): 825. PMID 20376121. doi:10.1038/464825b.
- ↑ «Briny deep basin may be home to animals thriving without oxygen». Science News. 23 de septiembre de 2013.
- ↑ Gould, 1989, pp. 124-136
- ↑ Bavestrello, Giorgio; Christian Sommer; Michele Sarà (1992). «Bi-directional conversion in Turritopsis nutricula (Hydrozoa)». Scientia Marina 56 (2-3): 137-140.
- ↑ Piraino, Stefano; F. Boero; B. Aeschbach; V. Schmid (1996). «Reversing the life cycle: medusae transforming into polyps and cell transdifferentiation in Turritopsis nutricula (Cnidaria, Hydrozoa)». Biological Bulletin (Biological Bulletin, vol. 190, no. 3) 190 (3): 302-312. JSTOR 1543022. doi:10.2307/1543022.
- ↑ Aurelia aurita Encyclopedia of Life. Retrieved 18 August 2016.
- ↑ Karleskint G, Richard Turner R and, James Small J (2012) Introduction to Marine Biology Cengage Learning, edition 4, page 445. ISBN 9781133364467.
- ↑ Holsinger, K. (2005). Keystone species. Retrieved 10 May 2010, from «Archived copy». Archivado desde el original el 30 de junio de 2010. Consultado el 12 de mayo de 2010.
- ↑ Black, Richard (26 de abril de 2008). «Colossal squid out of the freezer». BBC News.
- ↑ Ewen Callaway (2 de junio de 2008). «Simple-Minded Nautilus Shows Flash of Memory». New Scientist. Consultado el 7 de marzo de 2012.
- ↑ Kathryn Phillips (15 de junio de 2008). «Living Fossil Memories». Journal of Experimental Biology 211 (12): iii. doi:10.1242/jeb.020370.
- ↑ Robyn Crook & Jennifer Basil (2008). «A biphasic memory curve in the chambered nautilus, Nautilus pompilius L. (Cephalopoda: Nautiloidea)». Journal of Experimental Biology 211 (12): 1992-1998. PMID 18515730. doi:10.1242/jeb.018531.
- ↑ a b c Ruppert, Edward E.; Fox, Richard, S.; Barnes, Robert D. (2004). Invertebrate Zoology (7th edición). Cengage Learning. ISBN 81-315-0104-3.
- ↑ a b Hayward, PJ (1996). Handbook of the Marine Fauna of North-West Europe. Oxford University Press. ISBN 0-19-854055-8.
- ↑ Giribet, G.; Okusu, A, A.; Lindgren, A.R., A. R.; Huff, S.W., S. W.; Schrödl, M, M.; Nishiguchi, M.K., M. K. (May 2006). «Evidence for a clade composed of molluscs with serially repeated structures: monoplacophorans are related to chitons» (Free full text). Proceedings of the National Academy of Sciences of the United States of America 103 (20): 7723-7728. Bibcode:2006PNAS..103.7723G. PMC 1472512. PMID 16675549. doi:10.1073/pnas.0602578103.
- ↑ Healy, J.M. (2001). «The Mollusca». En Anderson, D.T., ed. Invertebrate Zoology (2nd edición). Oxford University Press. pp. 120-171. ISBN 0-19-551368-1.
- ↑ «Archived copy». Archivado desde el original el 26 de enero de 2011. Consultado el 10 de marzo de 2011.
- ↑ Braddy, Simon J.; Poschmann, Markus; Tetlie, O. Erik (2007). «Giant claw reveals the largest ever arthropod». Biology Letters 4 (1): 106-109. PMC 2412931. PMID 18029297. doi:10.1098/rsbl.2007.0491.
- ↑ Cressey, Daniel (21 de noviembre de 2007). «Giant sea scorpion discovered». Nature. doi:10.1038/news.2007.272. Consultado el 10 de junio de 2013.
- ↑ Patrick Kilday (28 de septiembre de 2005). «Mantis shrimp boasts most advanced eyes». The Daily Californian. Archivado desde el original el 29 de septiembre de 2012. Consultado el 7 de noviembre de 2018.
- ↑ S. N. Patek & R. L. Caldwell (2005). «Extreme impact and cavitation forces of a biological hammer: strike forces of the peacock shrimp». Journal of Experimental Biology 208 (19): 3655-3664. PMID 16169943. doi:10.1242/jeb.01831.
- ↑ D. R. Currie; T. M. Ward (2009). South Australian Giant Crab (Pseudocarcinus gigas) Fishery (PDF). South Australian Research and Development Institute. Fishery Assessment Report for PIRSA. Archivado desde el original el 28 de marzo de 2012. Consultado el 9 de diciembre de 2013.
- ↑ «An ugly giant crab of Japan». Popular Science 96 (6): 42. 1920.
- ↑ «FishBase: A Global Information System on Fishes». FishBase. Consultado el 17 de enero de 2017.
- ↑ «How Many Fish In The Sea? Census Of Marine Life Launches First Report». Science Daily. Consultado el 17 de enero de 2017.
- ↑ a b Wroe, S.; Huber, D. R.; Lowry, M.; McHenry, C.; Moreno, K.; Clausen, P.; Ferrara, T. L.; Cunningham, E.; Dean, M. N.; Summers, A. P. (2008). «Three-dimensional computer analysis of white shark jaw mechanics: how hard can a great white bite?». Journal of Zoology 276 (4): 336-342. doi:10.1111/j.1469-7998.2008.00494.x.
- ↑ Pimiento, Catalina; Dana J. Ehret; Bruce J. MacFadden; Gordon Hubbell (10 de mayo de 2010). «Ancient Nursery Area for the Extinct Giant Shark Megalodon from the Miocene of Panama». En Stepanova, Anna, ed. PLoS ONE (Panama: PLoS.org) 5 (5): e10552. Bibcode:2010PLoSO...510552P. PMC 2866656. PMID 20479893. doi:10.1371/journal.pone.0010552. Consultado el 12 de mayo de 2010.
- ↑ Lambert, Olivier; Bianucci, Giovanni; Post, Klaas; de Muizon, Christian; Salas-Gismondi, Rodolfo; Urbina, Mario; Reumer, Jelle (1 de julio de 2010). «The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru». Nature (Peru) 466 (7302): 105-108. Bibcode:2010Natur.466..105L. PMID 20596020. doi:10.1038/nature09067.
- ↑ Nielsen, Julius; Hedeholm, Rasmus B.; Heinemeier, Jan; Bushnell, Peter G.; Christiansen, Jørgen S.; Olsen, Jesper; Ramsey, Christopher Bronk; Brill, Richard W.; Simon, Malene; Steffensen, Kirstine F.; Steffensen, John F. (2016). «Eye lens radiocarbon reveals centuries of longevity in the Greenland shark (Somniosus microcephalus)». Science 353 (6300): 702-704. Bibcode:2016Sci...353..702N. PMID 27516602. doi:10.1126/science.aaf1703. Resumen divulgativo – Sci News (12 August 2016).
- ↑ Black, Richard (11 de junio de 2007). «Sawfish protection acquires teeth». BBC News.
- ↑ Marshall, A.; Bennett, M.B.; Kodja, G.; Hinojosa-Alvarez, S.; Galvan-Magana, F.; Harding, M.; Stevens, G.; Kashiwagi, T. (2011). «Manta birostris». The IUCN Red List of Threatened Species (IUCN) 2011: e.T198921A9108067. doi:10.2305/IUCN.UK.2011-2.RLTS.T198921A9108067.en. Archivado desde el original el 4 de abril de 2019. Consultado el 5 de noviembre de 2018.
- ↑ Lalli, C.; Parsons, T. (1993). Biological Oceanography: An Introduction. Butterworth-Heinemann. ISBN 0 7506 3384 0.
- ↑ John Dolan (November 2012). «Microzooplankton: the microscopic (micro) animals (zoo) of the plankton». Archivado desde el original el 4 de marzo de 2016. Consultado el 1 de abril de 2018.
- ↑ Harvey, Edmund Newton (1952). Bioluminescence. Academic Press.
Bibliografía editar
- Halpern, B.S.; Walbridge, S.; Selkoe, K.A.; Kappel, C.V.; Micheli, F.; D'Agrosa, C.; Bruno, J.F.; Casey, K.S.; Ebert, C.; Fox, H.E.; Fujita, R. (2008). «A global map of human impact on marine ecosystems». Science 319 (5865): 948-952. PMID 18276889. doi:10.1126/science.1149345.
- Paleczny, M.; Hammill, E.; Karpouzi, V.; Pauly, D. (2015). «Population trend of the world's monitored seabirds, 1950-2010». PLoS ONE 10 (6): e0129342. PMC 4461279. PMID 26058068. doi:10.1371/journal.pone.0129342.
- After 60 million years of extreme living, seabirds are crashing The Guardian, 22 September 2015.
- Ruppert, E.E.; Fox, R.S.; Barnes, R.D. (2004). Invertebrate Zoology (7th edición). Brooks / Cole. ISBN 0-03-025982-7.
 Datos: Q1230015
Datos: Q1230015