Myxogastrea
Myxogastrea o Myxogastria es el grupo principal de los mixomicetos, protistas conocidos comúnmente como mohos mucilaginosos.[2][3] Estos organismos toman tres formas distintas durante el transcurso de su vida: mixamebas unicelulares, una etapa de agregación denominada plasmodio y un cuerpo fructífero formador de esporas.[4][5] Myxogastrea se distingue de los otros grupos de mixomicetos por la formación de un verdadero plasmodio. Esto es, las mixamebas actúan como gametos fusionándose para dar lugar a un zigoto, cuyo núcleo se divide repetidamente y la célula se constituye en una gran masa protoplasmática y plurinucleada sin ningún tipo de pared, denominada plasmodio.[1] Los plasmodios se desplazan sobre el suelo ingiriendo todo tipo de materia orgánica y pueden llegar a cubrir varios metros cuadrados. El grupo comprende más de 900 especies conocidas.[6][7]
| Myxogastrea | ||
|---|---|---|
 Diderma testaceum | ||
| Taxonomía | ||
| Dominio: | Eukarya | |
| Reino: | Protista | |
| Filo: | Amoebozoa | |
| Subfilo: | Conosa | |
| Infrafilo: | Mycetozoa | |
| Clase: | Myxogastrea | |
| Órdenes y familias[1] | ||
| ||
Características
editarMyxogastrea es el grupo principal de Mycetozoa,[8][4][9][5] también conocido como Myxomycota, aunque ahora ya se sabe que no es de la familia de los hongos, clasificándose actualmente como protistas. El nombre latino proviene del griego myxo, que significa "barro", y mycota, que quiere decir hongo. Coloquialmente son conocidos como mohos plamodiales o acelulares del barro, o dentro de la comunidad científica como mixomicetos.
La clase se distribuye en todo el mundo, siendo más común en las regiones templadas donde tiene una biodiversidad mayor que en las regiones polares, los trópicos o subtrópicos. Se encuentran principalmente en los bosques abiertos, sobre troncos caídos, corteza de árboles vivos, suelo, hojarasca, estiércol y desechos. Aun así, también se encuentran en regiones extremas, tales como desiertos, bajo mantos de nieve o bajo el agua. Algunos viven sobre la corteza de los árboles, a veces en el dosel, y se conocen como mixomicetos corticícolas. El tamaño varía entre menos de 1 mm en las especies más pequeñas hasta llegar a alcanzar el metro cuadrado de superficie en las especies más grandes.
Estos organismos son exclusivamente heterotrofos, depredadores de marañas de comida microbiana, y tanto son capaces de envolver y digerir bacterias, levaduras, esporas de hongos, como materia en descomposición.[10][11]
- Distintas etapas del ciclo de vida de Myxogastrea
-
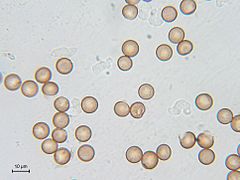 Esporas (esferas de color naranja) y mixamebas (en color blanco) de Symphytocarpus flaccidus
Esporas (esferas de color naranja) y mixamebas (en color blanco) de Symphytocarpus flaccidus -
 Physarum polycephalum (plasmodio)
Physarum polycephalum (plasmodio) -
 Fuligo septica (cuerpo fructífero).
Fuligo septica (cuerpo fructífero).


_(2188452157).jpg)
Ciclo de vida
editar
El ciclo de vida de estos organismos comprende las etapas de espora, mixameba, plasmodio y cuerpo fructífero.
La espora del mixomiceto es la fase de reposo y de dispersión de estos organismos.
Cuando germina una espora, se rompe, liberando una célula ameboide llamada mixameba. Dependiendo de las condiciones del entorno, la mixameba puede producir flagelos. Este proceso es reversible por lo que los flagelos se pueden retraer y la mixameba se establece de nuevo. La célula se alimenta mediante fagocitosis y se divide al tiempo que se alimenta.
Cuando dos mixamebas de diferentes poblaciones se encuentran, se fusionan formando un zigoto. El zigoto no se dividirá, pero en vez de eso crecerá al tiempo que su núcleo se divide, formando una larga, multinuclear célula llamada plasmodio. El plasmodio continuará consumiendo materia orgánica mediante fagocitosis.
Después de algún tiempo, por causas desconocidas, el plasmodio se convertirá en una estructura esporífera llamada cuerpo fructífero. Las esporas se separan del cuerpo fructífero y el ciclo de vida comienza de nuevo.
Esporas
editarLas esporas de Myxogastrea son haploides y miden usualmente entre 5 y 20 µm, alcanzando raramente 24 µm de diámetro. Su superficie es reticular, generalmente verrugosa o espinosa, aunque en raras ocasiones es lisa. El color, forma y diámetro de las esporas son características importantes para la identificación de especies. Los factores más importantes para la germinación de las esporas son la humedad y la temperatura. Las esporas pueden permanecer viables por varios años, dándose casos de esporas conservadas en herbarios que germinaron después de 75 años. Durante su formación, las esporan reciben un núcleo diploide que se convierte en haploide mediante meiosis. En la germinación, la cubierta de las esporas se abre a través de poros germinales especiales o bien de forma irregular, liberando de uno a cuatro protoplastos haploides.[12]
Mixamebas y mixoflagelados
editarDependiendo de las condiciones medioambientales, de las esporas brotan mixamebas o bien mixoflagelados. Las amebas se arrastran sobre el sustrato y se producen en condiciones secas. Los mixoflagelados pueden nadar y se desarrollan en ambientes húmedos. Casi siempre tienen dos flagelos, de los cuales generalmente uno es más corto que el otro y a veces vestigial. Los flagelos se utilizan para la locomoción y para ayudar a atraer a las partículas de alimentos. Si cambian las condiciones de humedad, las células pueden alternar entre los tipos de mixameba y mixoflagelado. Ninguna de las dos formas presenta pared celular.[13]
Esta etapa de desarrollo y también la siguiente son tróficas (fases de alimentación), en las cuales el organismo consume alimento y se desarrolla. Esta primera etapa trófica es unicelular y el organismo se alimenta de bacterias, esporas de hongos y desechos, y se reproduce únicamente a través de la división celular. Si las condiciones ambientales cambian adversamente en esta etapa, por ejemplo por temperaturas extremas, sequedad extrema o escasez de alimentos, la célula puede cambiar a un estado de reposo formando un microquiste. Para ello, la mixameba toma una forma esférica y segrega una pared celular delgada. En este estado puede sobrevivir fácilmente un año o más. Si las condiciones de vida mejoran, pasa al estado activo de nuevo.
Si dos células del mismo tipo se encuentran en esta fase, se fusionan formando un cigoto diploide mediante la fusión de protoplasmas y núcleos. Las condiciones que provocan esto no se conocen. El cigoto diploide se convierte en un plasmodio multinucleado a través de múltiples divisiones nucleares sin que tenga lugar la división celular. Si las células iniciales eran mixoflageladas, cambian su forma antes de la fusión a mixamebas. Para la producción de un cigoto, se requieren células de diferentes tipos, que en este caso se denominan "+" y "-", en vez de "masculino" y "femenino", puesto que estos organismos son heterotálicos.
Algunas especies de Myxogastrea pueden reproducirse asexualmente y son permanentemente diploides. No hay meiosis antes de la germinación de las esporas y los plasmodios se forman sin la fusión de dos células.
Plasmodio
editarLa segunda fase trófica comienza con el desarrollo del plasmodio. El organismo multinucleado ahora absorbe mediante fagocitosis tantos nutrientes como le sea posible. Estos son bacterias, protistas, sustancias disueltas, mohos, hongos superiores y pequeñas partículas de material orgánico. Esto permite que la célula tenga a un enorme crecimiento. El núcleo se divide múltiples veces y la célula pronto se hace visible a simple vista y por lo general y dependiendo de la especie, puede ocupar una superficie de hasta un metro cuadrado. Excepcionalmente, en 1987 se cultivó artificialmente un espécimen de Physarum polycephalum que alcanzó una superficie de 5,5 metros cuadrados. Las especies de Myxogastria tienen numerosos núcleos en su fase de plasmodio trófico; los más pequeños protoplasmodios no veteados puede tener entre 8 y 100 núcleos, mientras que las grandes mallas veteadas pueden tener entre 100 núcleos y 10 millones. Todos estos núcleos siguen siendo parte de una misma célula, que tiene una consistencia viscosa y puede ser transparente, blanca o de colores brillantes como naranja, amarillo o rosa.[14] Dependiendo del tamaño se distinguen tres tipos de plasmodios:
- Protoplasmodio. Es de tamaño microscópico, no forma venas y suele dar lugar a un solo esporangio.
- Afanoplasmodio. Es pequeño y difícil de ver a simple vista, forma una red de venas traslúcidas y da lugar a numerosos esporangios.
- Faneroplasmoido. De tamaño grande y visible, formando una red de grandes venas ramificadas en las que se puede observar el fluido que circula.
El plasmodio tiene capacidades de quimiotaxis positivas y negativas y fototaxis negativa, lo que significa que es capaz de moverse hacia los nutrientes y alejarse de las sustancias y luz peligrosas. Los movimientos se originan en el citoplasma granulado, que fluye en una dirección por pulsaciones dentro de la célula. De esta manera la célula alcanza una velocidad de hasta 1 mm por segundo. En esta fase puede producirse un estado de reposo, el llamado esclerocio, que es una forma endurecida compuesta por numerosos macroquistes, que permiten al organismo sobrevivir en esta fase a condiciones adversas, por ejemplo, durante los periodos de invierno o secos.
Cuerpo fructífero
editarLos plasmodios maduros pueden producir cuerpos fructíferos en las circunstancias adecuadas. Los factores exactos desencadenantes de este proceso son desconocidos. Según investigaciones de laboratorio en algunas especies puede deberse a cambios de humedad, temperatura o del valor del pH, así como de la escasez de alimento. El plasmodio abandona el desplazamiento y la ingesta de nutrientes y atraído por la luz (fototaxis positiva) se dirige hacia una zona de luz seca, para obtener una difusión óptima de las esporas. Una vez que comienza la fase de fructificación, esta no se puede detener. Si se producen perturbaciones, los cuerpos fructíferos a menudo producen esporas con malformaciones.
El plasmodio o partes de los cuerpos fructíferos pueden ser más pequeños que un milímetro, pero en casos extremos alcanzan hasta un metro cuadrado y pesan hasta 20 kilogramos (por ejemplo, Brefeldia maxima). Según la forma del cuerpo fructífero se distinguen los siguientes tipos:[15]
- Esporangio. Está formado por una esporoteca o bolsa que contiene la masa de esporas, que puede ser globosa, cilíndrica, etc. Además, la esporoteca puede estar situada sobre un pedúnculo o estipe o carecer de él.
- Plasmodiocarpo. Al igual que el anterior, contiene la masa de esporas, pero toma una forma veteada o mallada, que recuerda al plasmodio original.
- Etalio. Son grandes, con forma redondeada o de semiesfera, y con aspecto almohadillado, que contienen una gran masa de esporas.
- Seudoaetalio. De forma aparentemente similar al anterior, está formado por esporangios muy juntos pero que conservan su individualidad.
La esporoteca, que es lugar donde se encierra la masa de esporas, está protegida por una cubierta de capa simple o compuesta denominada peridio. El peridio eventualmente se abrirá para liberar las esporas, lo que puede hacerse por aberturas previamente predeterminadas o de forma irregular. Dentro de la esporoteca puede estar presente un capilicio, que es una estructura reticular o filamentosa que rodea a las esporas y les da soporte. Este se encuentra en casi todas las especies, excepto en Liceida y en las especies del género Echinostelium. Puede haber una columela, que es un eje que recorre longitudinalmente el esporocarpo y soporta la red del capilicio. Los cuerpos fructíferos casi siempre tienen un hipotalo en la base y en contacto con el sustrato. Cuando los cuerpos frutales abiertos se secan, las esporas son dispersadas por el viento o por animales pequeños, como cochinillas, ácaros o escarabajos, que o bien las recogen por contacto con los cuerpos fructíferos o bien las ingieren y luego los excretan. La dispersión por las corrientes de agua es también posible, pero juega un papel menor.
- Cuerpos fructíferos de Myxogastrea
-
 Esporangio (pediculado) de Trichia decipiens
Esporangio (pediculado) de Trichia decipiens -
 Plasmodiocarpo de Hemitrichia serpula
Plasmodiocarpo de Hemitrichia serpula -
 Etalio de Enteridium lycoperdon
Etalio de Enteridium lycoperdon -
 Seudoetalio de Tubifera ferruginosa
Seudoetalio de Tubifera ferruginosa
Macbr.jpg)

_lycoperdon,_Kyiv,_2012_09_20.jpg)

Clasificación
editarMyxogastrea se puede dividir en seis órdenes clasificados en dos subclases y dos superórdenes.[16] Los diferentes órdenes se pueden diferenciar atendiendo a los siguientes criterios:[17][18]
- Subclase Exosporeae. Comprende a los mixomicetos que forman tallos y esporas individualmente, esto es, las esporas se desarrollan externamente (de ahí el nombre de la subclase).
- Protosporangiida. Único orden de la subclase, considerado un grupo hermano de los demás, y anteriormente clasificado en la clase Protostelea.
- Subclase Myxogastria. Se caracterizan por presentar esporas que se desarrollan internamente y comprende a los restantes cinco órdenes.
- Superorden Lucisporidia. La masa de esporas es de coloración más o menos brillante. Dependiendo del orden, presentan o no un capilicio verdadero.
- Superorden Fuscisporidia o Columellidia. La masa de esporas suele ser de color oscuro (negro, marrón oscuro a marrón-rojizo), pero amarillo, hialino o débilmente coloreado sólo en Echinostelium y Semimorula (Echinosteliida). Además, presentan un capilicio verdadero.
- Echinosteliida. El cuerpo fructífero es pequeño, de menos de 0,5 mm de alto, en contraste con los siguientes dos grupos, en los que el cuerpo fructífero suele ser de mayor tamaño.
- Stemonitida. Carecen de carbonato cálcico en el cuerpo fructífero.
- Physarida. Contienen carbonato cálcico en alguna sección del cuerpo fructífero.
Filogenia
editarEl siguiente árbol filogenético muestra las relaciones entre los distintos grupos de mixomicetos.[4]
| Mycetozoa |
| ||||||||||||||||||||||||||||||||||||
Galería
editar







.jpg)

Véase también
editarReferencias
editar- ↑ a b Schnittler, M., Novozhilov, Y. K., Romeralo, M., Brown, M., & Spiegel, F. W. (2012). Fruit body-forming protists: Myxomycetes and myxomycete-like organisms. Englers Syllabus of Plant Families, 1, 1.
- ↑ Baldauf, S. L., & Doolittle, W. F. (1997). Origin and evolution of the slime molds (Mycetozoa). Proceedings of the National Academy of Sciences, 94(22), 12007-12012.
- ↑ Cavalier-Smith, T. (2013). Early evolution of eukaryote feeding modes, cell structural diversity, and classification of the protozoan phyla Loukozoa, Sulcozoa, and Choanozoa. European journal of protistology, 49(2), 115-178.
- ↑ a b c Fiore-Donno, A. M., Nikolaev, S. I., Nelson, M., Pawlowski, J., Cavalier-Smith, T., & Baldauf, S. L. (2010). Deep phylogeny and evolution of slime moulds (Mycetozoa). Protist, 161(1), 55-70.
- ↑ a b Baldauf SL, Doolittle WF (octubre de 1997). «Origin and evolution of the slime molds (Mycetozoa)». Proc. Natl. Acad. Sci. U.S.A. 94 (22): 12007-12. PMC 23686. PMID 9342353.
- ↑ Sina M. Ald et al. (2007) Diversity, Nomenclature, and Taxonomy of Protists, Syst. Biol. 56(4), 684–689, DOI: 10.1080/10635150701494127.
- ↑ Chapman, A. D. (2009). Numbers of living species in Australia and the world Archivado el 28 de septiembre de 2015 en Wayback Machine..
- ↑ «www.ncbi.nlm.nih.gov». Consultado el 27 de marzo de 2009.
- ↑ Fiore-Donno AM, Berney C, Pawlowski J, Baldauf SL (2005). «higher-order phylogeny of plasmodial slime molds (Myxogastria) based on elongation factor 1-A and small subunit rRNA gene sequences». J. Eukaryot. Microbiol. 52 (3): 201-10. PMID 15926995. doi:10.1111/j.1550-7408.2005.00032.x.
- ↑ C.J. Alexopolous, Charles W. Mims, M. Blackwell. 1996. Introductory Mycology, 4th ed. John Wiley and Sons, Hoboken NJ.
- ↑ Krishnan U, Barsamian A, Miller DL (2007). «Evolution of RNA editing sites in the mitochondrial small subunit rRNA of the Myxomycota». Meth. Enzymol. 424: 197-220. PMID 17662842. doi:10.1016/S0076-6879(07)24009-1.
- ↑ Wolfgang Nowotny: Myxomyceten (Schleimpilze) und Mycetozoa (Pilztiere) - Lebensformen zwischen Tier und Pflanze, 9783854740568, 45689126, German, English, French, Spanish.
- ↑ Steven L Stephenson; Henry Stempen: Myxomycetes : a handbook of slime molds, In: Myxomycetes, Timberland Press, isbn: 9780585342498, oclc: 47008442, pp. 15-18, 1994.
- ↑ Adl, S.M. et al. (2012). The revised classification of eukaryotes. Journal of Eukaryotic Microbiology, 59(5), 429-514.
- ↑ Microbiota Archivado el 26 de diciembre de 2015 en Wayback Machine.. (Consultado el 1/1/2016).
- ↑ Ruggiero, M. A., Gordon, D. P., Orrell, T. M., Bailly, N., Bourgoin, T., Brusca, R. C., Cavalier-Smith, T., Guiry, M.D. y Kirk, P. M. (2015). A Higher Level Classification of All Living Organisms.
- ↑ Una Introducción a los Myxomycetes (Consultado el 29/12/2015)
- ↑ Kretzschmar, M., Kuhnt, A., Bonkowski, M., & Fiore‐Donno, A. M. (2016). Phylogeny of the Highly Divergent Echinosteliales (Amoebozoa). Journal of Eukaryotic Microbiology.
Enlaces externos
editar Wikispecies tiene un artículo sobre Myxogastrea.
Wikispecies tiene un artículo sobre Myxogastrea. Wikimedia Commons alberga una categoría multimedia sobre Myxogastrea.
Wikimedia Commons alberga una categoría multimedia sobre Myxogastrea.
 Datos: Q46309
Datos: Q46309- Multimedia: Myxomycetes / Q46309
- Especies: Myxogastria